西太平洋沉积物中生物扰动及其对
有机碳输送的210Pb示踪
杨伟锋1,2,张馨星1,陈敏1,2,王宙1,王春生3,黄奕普1,2,张东声3,邱雨生1,2
(1. 厦门大学 海洋与环境学院,福建 厦门,361005;
2. 厦门大学 近海海洋环境科学国家重点实验室,福建 厦门,361005;
3. 国家海洋局第二海洋研究所,浙江 杭州,310012)
摘要:采用低本底g能谱仪测定了西太平洋海山区3个沉积物岩心浅层的210Pb和226Ra比活度。结果表明,表层10 cm以内210Pb均表现出相对于母体226Ra的过剩现象,反映了生物扰动作用的存在。基于过剩210Pb(210Pbex)的一维稳态模型得到沉积物的生物扰动系数Db介于1.59~8.64 cm2/a之间,平均为4.45 cm2/a,表征西太平洋海山区较强的生物扰动作用。各站位有机碳(TOC)含量与210Pbex比活度之间存在良好线性正相关关系,揭示了TOC含量的高低在很大程度上调控着生物扰动作用的强度。基于此关系,建立了生物扰动从表层向深层沉积物输送有机碳的一维模型,计算得3个站位总有机碳输送通量的上限值介于0.04~0.16 mmol/(cm2・a)之间,平均为0.10 mmol/(cm2・a)。
关键词:210Pb;生物扰动;POC;太平洋
中图分类号:P736.21 文献标志码:A 文章编号:1672-7207(2011)S2-0189-07
Bioturbation in the western Pacific Ocean: Use of 210Pb to evaluate intensity and POC transport
YANG Wei-feng1, 2, ZHANG Xin-xing1, CHEN Min1, 2, WANG Zhou1,
WANG Chun-sheng3, HUANG Yi-pu1, 2, ZHANG Dong-sheng3, QIU Yu-sheng1, 2
(1. College of Oceanography and Environmental Science, Xiamen University, Xiamen 361005, China;
2. State Key Laboratory of Marine Environmental Science, Xiemen University, Xiamen 361005, China;
3. The Second Institute of Oceanography, State Oceanic Administration, Hangzhou 310012, China)
Abstract: The specific activities of 210Pb and 226Ra in three sediment cores from the western Pacific were determined by γ spectrometry. Results show that bioturbation effect was revealed by the excess 210Pb (i.e. 210Pbex) relevant to it parent of 226Ra within the upper 10 cm. The bioturbation coefficient Db, evaluated from a one-dimensional steady-state model, varies from 1.59 cm2/a to 8.64 cm2/a with an average of 4.45 cm2/a. Positive correlations between 210Pbex and total organic carbon (TOC) were observed in three sediment cores, which indicates that the bioturbation in the western Pacific Ocean might be controlled by TOC content. A one-dimensional model is proposed to estimate the organic carbon transport induced by bioturbation based on these correlations. The upper threshold of fluxes of organic carbon from surface to subsurface sediment varies from 0.04 mmol/(cm2・a) to 0.16 mmol/(cm2・a) with the mean of 0.10 mmol/(cm2・a) from 3 sample stations.
Key words: 210Pb; bioturbation; POC; the Pacific Ocean
生物扰动是底栖生物通过摄食和掘穴等活动对沉积物进行混合的过程[1],全球海底沉积物生物扰动的平均深度为(9.8±4.5) cm[2]。生物扰动在一定程度上改变了沉积物的物理、化学和生物学特征[3-4],进而影响沉积物的早期成岩过程并改变沉积记录[5]。在早期成岩过程中,生物扰动作用影响着有机质的再循环和埋藏[6]、方解石等矿物的溶解[5],因此,在不同程度上平滑甚至扭曲了沉积物或矿物记录的古海洋学信息。通常用生物扰动系数来表征生物扰动作用[7]。生物扰动系数也是深海沉积物有机质再循环模型及古海洋学重建模型中的重要参数[7-8]。目前,计算生物扰动系数的方法有210Pb过剩法[9-11]、234Th过剩法[12-14]和Chl.a法[15]。
热带太平洋海底具有丰富的多金属矿物[16-17],记录了丰富的古海洋学信息。研究该海域生物扰动作用对成矿过程的影响有利于准确揭示该海域沉积物及矿物蕴藏的古海洋学记录。同时,研究生物扰动向深层沉积物输送的有机碳有利于定量了解深海底栖生态系统的物质流动和生态环境,对准确评价矿物开采可能引起的环境效应提供基线参数。但过去的研究多集中在中太平洋和东太平洋[9,12,18-19],而对西太平洋相关研究很少。由于采样等条件的限制,我国对深海生物扰动作用的研究尚非常少。李凤业等[20]研究了过冲绳海槽沉积物的生物混合作用,杨群慧等[18-19]应用210Pb过剩法共研究了东太平洋3个沉积物岩心的生物扰动作用。在西太平洋中国多金属合同区尚未见生物扰动作用的研究报道。本文作者采用210Pb示踪西太平洋 海山区生物扰动作用,并探索应用210Pb计算生物扰动对有机碳的向下输送通量。
1 样品采集与分析方法
1.1 样品采集及前处理
3个沉积物岩心(WPMC0801,WPMC0802和WPMC0803)由“大洋一号”考察船于2008年在西太平洋热带海域采用多管采样器采集(如图1所示)。采集后沉积物上覆水用虹吸去除,之后在船上实验室内将表层样品(6~10 cm以浅)按1 cm间隔垂直分割,分割后的子样装入洁净塑料袋中,于-20 ℃冷冻带回陆地实验室。在陆地实验室内将沉积物子样室温密封解冻,测量其质量,然后于100 ℃烘干至恒质量,据烘干前后质量差计算沉积物含水量及沉积物湿密度。取上述处理好的干沉积物样品1 g用于总有机质(TOM)的测量,剩余部分磨细、混匀并密封于70 mm?50 mm的聚乙烯g能谱测量样品盒内等待进行210Pb和226Ra的测量。
1.2 210Pb和226Ra测量
预处理好的沉积物样品在样品盒内密封25 d以上,以确保样品中226Ra与其测量的子体基本平衡。将待测样品置于EG&G Ortec公司生产的高纯锗低本底g能谱仪上测量,要求感兴趣峰面积误差小于10%。210Pb采用46.5 keV(4.0%)g射线峰,226Ra采用其子体214Pb在351.9 keV(37.09%)和214Bi在609.3 keV (46.1%)和1 120.3 keV(15.0%)的g射线峰。仪器的探测效率刻度及活度计算详见文献[21-22]。文中所给210Pb和226Ra数据均为单位质量干沉积物中的放射性活度,并校正至采样时刻。过剩210Pb(即210Pbex)比活度APbex为210Pb比活度APb减去其母体226Ra的比活度ARa。

图1 西太平洋沉积物采样站位图
Fig.1 Sampling stations in the western Pacific Ocean
1.3 TOM测量
取烘干后的沉积物1 g置于经500 ℃灼烧过8 h的陶瓷干锅内,于450 ℃下在马弗炉内灼烧至恒质量,称量灼烧前后质量差以计算沉积物中TOM含量。为了便于讨论,将总TOM含量转化为总有机碳(TOC)含量,转化系数为2[23]。根据文中计算生物扰动对TOC输送通量的需要,通过TOC含量和湿沉积物密度(ρ)将TOC含量转化为TOC的湿密度,即单位体积湿沉积物中的TOC含量,表示为CTOC(mmol/cm3)。
2 结果
2.1 210Pb和226Ra的分布
图2给出了3个站位沉积物中210Pb比活度,226Ra比活度和CTOC的分布。3个站位210Pb比活度APb接近,表现出从表层向下逐渐衰减的特征,类似的分布在热带太平洋海域都有报道[9,18-19]。通常认为深海沉积物中210Pb主要来自母体226Ra的衰变产生,若没有其他过程,APb的分布应类似于ARa的分布,但研究结果显示APb和ARa在WPMC0801和WPMC0802站的分布相反,该2个站位ARa随深度增加而增大,APb则随深度增加而减小(如图2所示),该差异表明210Pb不完全来自沉积物中226Ra的产生。同时,3个站位210Pb相对于226Ra都存在显著的过剩(表示为210Pbex)现象。WPMC0801,WPMC0802和WPMC0803站210Pbex的比活度APbex分别介于394~1 028 Bq/kg,517~833 Bq/kg和533~874 Bq/kg之间,平均分别为703 Bq/kg,671 Bq/kg和619 Bq/kg。显著过剩的210Pb进一步证实表层沉积物中其它来源的210Pb。结合210Pb的强颗粒活性,过剩210Pb应来自底层水中颗粒物清除的210Pb,该清除将导致底层水中210Pb相对于226Ra亏损,太平洋底层水中升高的颗粒态210Pb及加强的210Po相对于母体210Pb亏损[24]都支持底层水中较强的颗粒物清除210Pb过程。须指出的是:尽管底层水中清除的210Pb沉降在沉积物表面,但表面以下至10 cm深度,依然观测到210Pb明显过剩(如图2所示)。研究海域附近沉积物的沉积速率低于1 cm/ka[24],如没有混合过程,以210Pb仅22.3 a的半衰期其应在表层1 mm内与226Ra达到平衡,因此,图2中上层10 cm沉积物以内过剩210Pb的存在揭示了上层沉积物进行了明显的混合过程,该混合由底栖生物活动所致[9],这为研究海域210Pb过剩法示踪生物扰动过程奠定了基础。
2.2 TOC的分布
总体上,WPMC0801,WPMC0802和WPMC08033个站位CTOC的密度变化范围及平均值都比较接近,其平均值分别为0.48,0.45和0.49 mmol/cm3。CTOC分布表现为从表层向下减小的特征(如图2所示),说明表面沉积物具有高的TOC含量。在生物对沉积物进行混合扰动过程中,TOC会同时被混合至深层沉积物。APbex与CTOC之间的关系(图3)证实,由生物扰动的相关关系,据此关系,可以应用210Pbex法估算生物导致的表面以下210Pb的过剩程度与CTOC之间存在良好混合向深层沉积物输送的TOC通量。
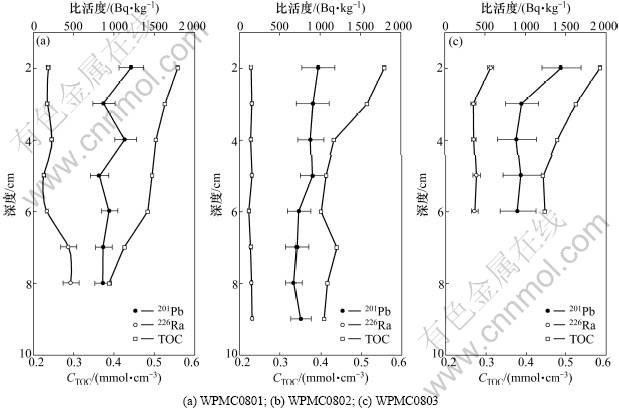
图2 沉积物中210Pb比活度,226Ra比活度和CTOC的分布
Fig.2 Profiles of 210Pb activity, 226Ra activity and CTOC in the western Pacific sediments
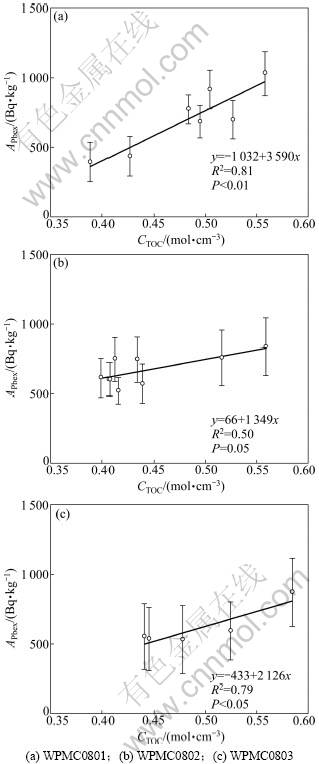
图3 沉积物中APbex与CTOC之相关关系
Fig.3 Relationship between specific activity of excess 210Pb and CTOC
3 讨论
3.1 生物扰动系数
生物扰动作用通常采用生物扰动系数进行量化,该系数通过Nozaki等[25]的一维模式计算:
 (1)
(1)
其中:z为深度(cm);APbex为深度z处210Pbex比活度(Bq/kg);t为时间(a);r代表沉积物密度(g/cm3);Db为生物扰动系数(cm2/a);s代表沉积速率(cm/a);l为210Pb的衰变常数(0.031 a-1)。
该模式将生物扰动过程视为类涡动扩散混合过程。假定沉积物处于稳定状态,沉积速率s在210Pb所示踪的时间尺度内(100 a)的影响可忽略,应用边界条件z=0 cm,APbex= APbex0和z?μ,APbex ?0,可解得式(1)中的
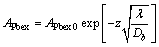 (2)
(2)
式中:APbex0为沉积物-水界面处APbex(Bq/kg)。由式(2)可见:APbex随深度应呈指数衰减,通过拟合210Pbex分布的指数方程可以获得生物扰动系数Db。
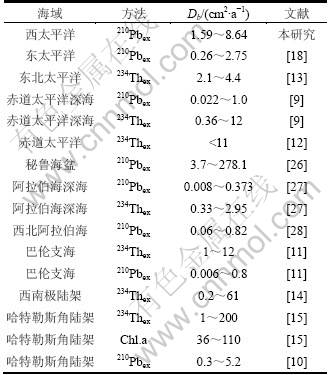
从图4可见:210Pbex与深度之间存在良好的相关性,证实上述模式适合于研究海域生物扰动系数的计算。WPMC0801,WPMC0802和WPMC0803站的Db分别为1.59,8.64和3.11 cm2/a,3站Db值平均为4.45 cm2/a。该结果与东太平洋和赤道太平洋深海的报道结果比较接近(表1),明显低于高生产力的秘鲁海盆和陆架/坡海域测值(表1),上述差异揭示了生物扰动作用的强度与海域的生产力之间可能存在联系,一些研究也已表明水柱的POC输出通量调控着沉积物混合层的深度和生物扰动的强度[2,7,9]。
3.2 生物扰动与TOC的关系
通常认为影响生物扰动的因素包括沉积物累积速率[29-30]、水深[31]、氧的穿透深度[7]和水柱POC的输出通量[7,9]。但在具体海域生物扰动作用的强度通常受主要影响因素调控。在本研究的西太平洋海山区,生物扰动作用可能受TOC含量的控制,该推测由下列证据支持:(1)图3中APbex与CTOC含量之间的正相关关系说明深层沉积物中TOC含量越高,从表层沉积物向下输送的过剩210Pb越多,即TOC促进了生物扰动作用;(2)研究表明距研究海域较近的热带中太平洋水柱POC的输出通量控制着底栖生物和化学过程的速 率[9,12],东太平洋多金属结核区的研究结果也表明有机碳含量越高,生物扰动作用越强[18];(3)Smith等[7]对三大洋深海36个站位的研究表明:水柱POC的输出通量通过控制底栖生物的大小和活动进而调节混合层的厚度和生物扰动的强度。由此可见:研究海域沉积物中有机碳的含量可能是生物扰动作用调控的主因素。但需指出的是:我们的研究站位比较少,量化有机碳含量对西太平洋沉积物中生物扰动作用的调控程度尚需更多研究。

图4 APbex随深度变化及生物扰动系数的计算
Fig.4 Variations of excess of 210Pb with depth and computation of bioturbation coefficients
表1 不同海域沉积物生物扰动系数
Table 1 Bioturbation coefficients in various ocean environments
尽管研究的3个站位生物扰动作用均主要受TOC含量调控,但TOC的影响程度并不相同。图3显示:APbex对CTOC的依赖程度差异较大,Db最大的WPMC0802站斜率最小,而Db最小的WPMC0801站斜率最大。该差异可能是不同站位沉积物中生物群落构成不同,而不同的生物对有机碳的需求不同所致。东太平洋结核区的研究发现合同区东、西亚区沉积物CTOC差异不足30%情况下生物扰动系数差异可达1个数量级,而这2个亚区的底栖生物结构差异显著,如多毛类比例差异近30%[18]。由此可见,研究海域生物扰动作用在TOC主要控制下,同时受生物组成等其他因素影响。
3.3 生物扰动对TOC的输送
生物扰动混合沉积物过程中会向深层沉积物输送有机质,为深层生物提供有机碳。根据APbex的分布模式及APbex与CTOC之间的关系,可以建立表面TOC向下输送的一维模型:
 (3)
(3)
式中:Fb代表TOC的通量(mmol/(cm2・a)),d(CTOC)/dz为CTOC的垂向密度梯度,负号表示通量方向向下。对WPMC0801,WPMC0802和WPMC0803 3个站位沉积物中CTOC的分布进行拟合,得到的方程分别为:
CTOC=0.614-0.026z (R2=0.94,P<0.01) (4)
CTOC=0.550-0.019z (R2=0.62,P<0.03) (5)
CTOC=0.640-0.036z (R2=0.90,P<0.02) (6)
由式(4)~(6)可见:研究站位CTOC与深度之间存在良好的线性正相关关系,即符合如下形式的方程:
CTOC=A+B?z (7)
求CTOC对深度z的微分,可得:
dCTOC/dz=B (8)
将式(4)~(6)中拟合的B和式(2)得到的生物扰动系数代入式(3),计算得WPMC0801,WPMC0802和WPMC0803站由生物扰动向下输送的TOC通量分别为0.04,0.16和0.11 mmol/(cm2・a),平均为0.10 mmol/(cm2・a)(表2)。
表2 CTOC垂向浓度梯度、生物扰动系数和TOC输送通量
Table 2 Concentration gradient of TOC, bioturbation coefficients and fluxes of TOC

需要指出的是:式(3)主要是基于生物扰动导致的深层沉积物中APbex与CTOC之关系来计算扰动过程对TOC的输送,虽然模型中的Db已经计入了210Pb向下混合过程中的衰变,但式(3)并未考虑此过程中TOC的降解,因此,该模型得到的结果代表了TOC输送的上限值。
4 结论
热带西太平洋沉积物中生物扰动系数(Db)与中部和东部热带太平洋接近,低于一些高生产力的上升流区和陆架/坡区。生物扰动作用主要受沉积物中TOC含量的调控,但同时受生物群落构成等因素的影响。通过生物扰动导致的深层沉积物中APbex与CTOC之间的关系可以量化生物活动从表层沉积物向深层输送的有机碳,该有机碳代表输送通量的上限值。本研究结果说明,210Pb可用于定量深海沉积物中生物扰动作用对底栖生态系统的物质流动及早期成岩过程的重要参数。
致谢:
感谢“大洋一号”科学考察船全体船员及相关单位同行在样品采集过程中给予的大力支持。
参考文献:
[1] Jumars P L, Mayer M, Deming J W, et al. Deep-Sea deposit-feeding strategies suggested by environmental and feeding constraints[J]. Philosophical Transactions of The Royal Society London, 1990, A331(1616): 85-101.
[2] Boudreau B P. Mean mixed depth of sediments: The wherefore and the why[J]. Limnology and Oceanography, 1998, 43(3): 524-526.
[3] Wheatcroft R A, Jumars P A, Smith C R, et al. A mechanistic view of the particulate biodiffusion coefficient: Step lengths, rest periods and transport directions[J]. Journal of Marine Research, 1990, 48(1): 177-207.
[4] Aller R C. Bioturbation and remineralization of sedimentary organic matter: Effects of redox oscillation[J]. Chemical Geology, 1994, 114(3/4): 331-345.
[5] Henderson G M, Lindsay F N, Slowey N C. Variation in bioturbation with water depth on marine slopes a study on the Little Bahamas Bank[J]. Marine Geology, 1999, 160(1/2): 105-118.
[6] Smith C R, Pope r H, Demaster D J, et al. Age-dependent mixing of deep-sea sediments[J]. Geochimica et Cosmochimica Acta, 1993, 57(7): 1473-1488.
[7] Smith C R, Rabouille C. What controls the mixed-layer depth in deep-sea sediments? The importance of POC flux[J]. Limnology and Oceanography, 2002, 49(2): 418-426.
[8] Rabouille C, Gaillard J F, Tréguer P, et al. Biogenic silica recycling in surficial sediments across the Polar Front of the Southern Ocean (Indian Sector)[J]. Deep-Sea Research II, 1997, 44(5): 1151-1176.
[9] Smith C R, Berelson W, Demaster D J, et al. Latitudinal variations in benthic processes in the abyssal equatorial Pacific: control by biogenic particle flux[J]. Deep-Sea Research II, 1997, 44(9/10): 2295-2317.
[10] Alperin M J, Suayah I B, Benninger L K, et al. Modern organic carbon burial fluxes, recent sedimentation rates, and particle mixing rates from the upper continental slope near Cape Hatteras, North Carolina (USA)[J]. Deep-Sea Research II, 2002, 49(20): 2645-4665.
[11] Carroll J, Zaborska A, Papucci C, et al. Accumulation of organic carbon in western Barents Sea sediments[J]. Deep-Sea Research II, 2008, 55(20/21): 2361-2371.
[12] Pope R H, Demaster D J, Smith C R, et al. Rapid bioturbation in equatorial Pacific sediments: Evidence from excess 234Th measurements[J]. Deep-Sea Research II, 1996, 43(4/6): 1339-1364.
[13] Smith C R, Maybaum H L, Baco A R, et al. Sediment community structure around a whale skeleton in the deep Northeast Pacific: Macrofaunal, microbial and bioturbation effects[J]. Deep-Sea Research II, 1998, 45(1/3): 335-364.
[14] Mcclintic M A, Demaster D J, Thomas C J, et al. Testing the FOODBANCS hypothesis: Seasonal variations in near-bottom particle flux, bioturbation intensity, and deposit feeding based on 234Th measurements[J]. Deep-Sea Research II, 2008, 55(22/23): 2425-2437.
[15] Green M A, Aller R C, Cochran J K, et al. Bioturbation in shelf/slope sediments off Cape Hatteras, North Carolina: the use of particle and solute transport[J]. Deep-Sea Research II, 2002, 49(20): 4627-4644.
[16] 韩喜球, 李家彪, 王英, 等. 分形理论在太平洋多金属结核分布研究中应用[J]. 海洋学报, 2001, 23(3): 142-146.
HAN Xi-qiu, LI Jia-biao, WANG Ying, et al. Application of fractal theory to distribution of polymetallic nodules from the Pacific Ocean[J]. Acta Oceanologica Sinica, 2001, 23(3): 142-146.
[17] Veillette J, Sarrazin J, Gooday A J, et al. Ferromanganese nodule fauna in the Tropical North Pacific Ocean: Species richness, faunal cover and spatial distribution[J]. Deep-Sea Research I, 2007, 54(11): 1912-1935.
[18] 杨群慧, 周怀阳. 中国多金属结核合同区近表层沉积物生物扰动作用的过剩210Pb证据[J]. 科学通报, 2004, 49(21): 2198-2203.
YANG Qun-hui, ZHOU Huai-yang. Bioturbation in near-surface sediments from the COMRA Polymetallic Nodule Area: Evidence from excess 210Pb measurements[J]. Chinese Science Bulletin, 2004, 49(21): 2198-2203.
[19] 杨俊鸿. 北太平洋与同安湾若干海洋学过程的同位素示踪[D]. 厦门: 厦门大学海洋与环境学院, 2007: 57-84.
YANG Jun-hong. Isotope tracers for marine processes in the North Pacific and the TongAn Bay[D]. Xiamen: Xiamen University. College of Oceanography and Environmental Science, 2007: 57-84.
[20] 李凤业, 谭长伟, 史玉兰, 等. 冲绳海槽沉积物混合作用的研究[J]. 海洋科学, 1996, 6: 57-84.
LI Feng-ye, TAN Chang-wei, SHI Yu-zhu, et al. Mixing rate of sediment in the Okinowa Trough[J]. Marine Sciences, 1996, 6: 57-84.
[21] 刘广山, 黄奕普. 沉积物中238U等9种放射性核素g谱法同时测定[J]. 台湾海峡, 1998, 17(4): 359-363.
LIU Guang-shan, HUANG Yi-pu. Simultaneous measurement of 238U, 235U, 210Pb, 228Th, 228Ra, 226Ra, 40K, 60Co and 137Cs in sediment samples using HPGe γ spectrometer[J]. Journal of Oceanography in Taiwan Strait, 1998, 17(4): 359-363.
[22] 刘广山, 陈敏, 黄奕普, 等. 海洋沉积物岩心放射性核素的g谱测定[J]. 厦门大学学报: 自然科学版, 2001, 40(3): 669-674.
LIU Guang-shan, CHEN Min, HUANG Yi-pu, et al. Measurement of radionuclides in sediments core by HPGe γ spectroscopy[J]. Journal of Xiamen University: Natural Science, 2001, 40(3): 669-674.
[23] Chase Z, Anderson R F, Fleisher M Q, et al. The influence of particle composition and particle flux on scavenging of Th, Pa and Be in the ocean[J]. Earth and Planetary Science Letters, 2002, 204(1/2): 215-219.
[24] 库兹涅佐夫著, 海洋放射年代学[M], (1976). 夏明等译. 科学出版社, 1980: 1-41.
KUZNETSOV. Marine Radiochronology[M], (1976). XIA Ming et al. Beijing: Science Press, 1980: 1-41.
[25] Nozaki Y, Cochran J K, Turekian K K, et al. Radiocarbon and 210Pb distribution in submersible deep-sea cores from Project FAMOUS[J]. Earth and Planetary Science Letters, 1977, 34(2): 167-173.
[26] Suchow A, Treppke U, Wiedicke M H, et al. Bioturbation coefficients of deep-sea sediments from the Peru Basin determined by gamma spectrometry of 210Pbex[J]. Deep-Sea Research II, 2001, 48(17/18): 3569-3592.
[27] Turnewitsch R, Witte U, Graf G. Bioturbation in the abyssal Arabian Sea: Influence of fauna and food supply[J]. Deep-Sea Research II, 2000, 47(14): 2877-2911.
[28] Smith C R, Levin L A, Hoover D J, et al. Variations in bioturbation across the oxygen minimum zone in the northwest Arabian Sea[J]. Deep-Sea Research II, 2000, 47(1/2): 227-257.
[29] Boudreau B P. Is burial velocity a master parameter for bioturbation?[J]. Geochimica et Cosmochimica Acta, 1994, 58(4): 1243-1249.
[30] Tromp T K, Cappellen P V, Key R M, et al. A global model for the early diagenesis of organic carbon and organic phosphorus in marine sediments[J]. Geochimica et Cosmochimica Acta, 1995, 59(7): 1259-1284.
[31] Middelburg J J, Soetaert K, Herman P M J. Empirical relationships for use in global diagenetic models[J]. Deep-Sea Research, 1997, 44(2): 327-344.
(编辑 邓履翔)
收稿日期:2011-06-15;修回日期:2011-07-15
基金项目:中国大洋协会“十一五”课题(DYXM-115-01-3-04, DYXM-115-02-4-06);国家自然科学基金资助项目(41076043, 40606022);福建省自然科学基金资助项目(2009J05105)
通信作者:杨伟锋(1978-),男,山西晋城人,博士,副教授,从事同位素海洋学研究,电话:0592-2189468;E-mail: wyang@xmu.edu.cn

