DOI:10.19476/j.ysxb.1004.0609.2019.03.21
Leptospirillum ferriphilum耐砷能力的驯化及驯化机制
郑兴福1,聂珍媛1,夏金兰1,刘李柱1,杨洪英2
(1. 中南大学 资源加工与生物工程学院,教育部生物冶金重点实验室,长沙 410083;
2. 东北大学 冶金学院,沈阳 110819)
摘 要:为了获得耐砷能力较高的中度嗜热嗜酸浸矿菌Leptospirillum ferriphilum,采用连续转接驯化法进行耐砷驯化,并对菌株的驯化机制进行研究。结果表明:驯化后Leptospririllum ferriphilum菌的As(Ⅲ)和As(Ⅴ)耐受浓度分别提高到7.0 g/L和11.0g/L;As(Ⅴ)驯化菌株对砷黄铁矿的浸出作用明显比As(Ⅲ)驯化菌株强;砷驯化菌细胞官能团发生明显变化,多糖和蛋白质含量明显增多,细胞表面负电荷明显降低,以适应高砷的环境。同步辐射原位C-K边XANES谱图表明,驯化菌糖类和蛋白质的分布位置发生了变化;DFT理论计算表明,驯化菌更容易吸附到矿物表面上。
关键词:砷;砷黄铁矿;黄铁矿;驯化
文章编号:1004-0609(2019)-03-0617-11 中图分类号:TF18 文献标志码:A
在含砷金矿的冶炼工艺中,常用浸矿微生物预氧化含砷金矿,其中浸矿菌通过自身代谢作用,浸出含砷金矿中的砷、铁与硫,从而分离出金矿。在氧化预处理过程中,释放的砷在溶液中累积;当溶液中的可溶性砷累积到一定浓度时,浸矿菌的生长及浸出活性受到抑制,预氧化处理效率降低;为了保持浸矿菌活性,工艺中必须每隔一定时间更换或稀释高砷浓度的溶液,这大大增加劳动强度与生产成本[1]。在工业生产中,浸矿菌预处理含砷金矿时,砷含量大多控制在2%~6%(质量分数)之间[2];自然界中的原矿或次生矿,含砷量超过6%。提高浸矿菌抗砷能力,是有效氧化预处理这类含砷金矿的前提,因此,对所得的浸矿菌进行砷的驯化,提高浸矿菌对砷的耐受能力在工业生产中具有极其重要的意义。
已有研究者将Acidithiobacillus ferrooxidans的耐砷能力提高至6 g/L[3],将Acidithiobacillus caldus的耐砷能力提高至3.38 g/L[4],将Leptospirillum ferriphilum (L. ferriphilum)的耐砷能力驯化至4.88 g/L[5];吴俊子等[6]通过对中度嗜热混合菌的驯化,成功培养出耐受矿浆质量浓度为200 g/L、砷质量浓度8 g/L的高耐砷混合菌。有研究指出砷驯化菌与原始菌相比,蛋白质的表达量和表达类型均存在差异[7];溶液中亚砷酸盐或砷酸盐浓度的不同,会诱导微生物细胞内的砷抗性蛋白种类和表达量出现差异[8]。富瑶等[9]指出,As(Ⅲ)胁迫时胞外多糖含量增长超过34.14%,As(Ⅴ)胁迫时胞外多糖含量增长超过28.84%。目前所得到的驯化菌耐砷能力还不足以应对生物浸出过程的高浓度砷环境,其耐砷驯化机制也还需要进一步的探讨。本实验通过逐渐加大砷浓度驯化获得菌株,期望得到耐砷性强的菌株,并阐释菌株对高砷环境的适应机制,为后期开展含砷金矿矿物的预处理及高浓度砷污染环境的治理提供依据。
L. ferriphilum为中度嗜热嗜酸的好氧革兰氏阴性菌,是一种代表性的浸矿微生物。本文作者以L. ferriphilum菌为研究对象,以黄铁矿作为能源底物,采用连续转接驯化法[3]进行耐砷驯化,在不同浓度的As(Ⅲ)和As(Ⅴ)条件下进行耐砷驯化,以期获得耐砷能力较高的驯化菌株,并进一步对菌株的驯化耐砷机制进行探讨。
1 实验
1.1 菌种及培养基
L. ferriphilum菌来源于中南大学生物冶金重点实验室。摇瓶培养条件如下:pH 2.0,温度 45 ℃,转速170 r/min。采用 MSN 培养基,配方如下:(NH4)2SO4 3.0 g/L,Na2SO4 1.41 g/L,KCl 0.1 g/L,K2HPO4 0.05 g/L,MgSO4・7H2O 5.0 g/L,Ca(NO3)2 0.1 g/L,FeCl3・6H2O 1.1×10-3 g/L,CuSO4 5.0×10-5 g/L,HBO3 2.0×10-4 g/L,MnSO4・H2O 8.0×10-5 g/L,CoCl・H2O 6.0×10-5 g/L,ZnSO4・7H2O 9.0×10-5 g/L,使用硫酸调节pH 2.0。
1.2 菌株的耐砷性驯化
本实验中As(Ⅲ)溶液用亚砷酸钠配制,As(Ⅴ)溶液用五氧化二砷(As2O5)配制。
1.2.1 砷的胁迫实验
对L. ferriphilum菌设置As(Ⅲ)胁迫浓度梯度:0 g/L,1.0 g/L,2.0 g/L,3.0 g/L,4.0 g/L,5.0 g/L;设置As(Ⅴ)浓度梯度:0 g/L,2.0 g/L,4.0 g/L,6.0 g/L,8.0 g/L,9.0 g/L;初始接种量均为8×107 mL-1,每隔12 h取样一次,使用显微镜观察细菌浓度变化情况,确定砷对菌的胁迫浓度。
1.2.2 菌株的驯化
菌株以黄铁矿为能源,按8×107 cell/mL的接种量接种到含As(Ⅲ)或As(Ⅴ)的培养基中进行驯化,当细菌生长繁殖至稳定期时,砷浓度按照As(Ⅲ) 1 g/L,As(Ⅴ) 2 g/L的浓度梯度递增进行下一轮的驯化。
1.2.3 砷黄铁矿的浸出实验
将驯化菌与原始菌接种到含有砷黄铁矿的培养基体系中,接种量为4×107 mL-1,测定相关浸矿参数。
在驯化过程中,分别以细菌计数板、PHS-3C型pH计和PHS-25型数显酸度计(以Ag/AgCl为参比电极)测定菌株在生长过程悬浮液中细菌的浓度、pH和氧化还原电位的变化[10];以磺基水杨酸法[10]和电感耦合等离子体发射光谱仪(ICP-OES)分别测定溶液中铁离子和砷离子的浓度变化;以扫描电镜(SEM,美国 FEI公司)表征驯化前后细菌对矿物的作用。
1.3 驯化机制
驯化完成后,选取细胞组分差距最大的驯化菌和原始菌进行更进一步的驯化机制分析。
1.3.1 糖类与蛋白质的测量
以红外吸收光谱[11] (Nicolet Nexus 670, USA)、荧光显微镜(德国莱卡公司 DM2500)、BCA蛋白质检测试剂盒和硫酸法测量驯化前后菌株糖类和蛋白质的变化[12]。
1.3.2 驯化前后L. ferriphilum菌表面性质的探究
以动电位观察菌株驯化前后细胞表面电位的变化;以同步辐射C-K边X射线吸收近边结构光谱(上海同步辐射装置)分析菌株驯化前后细胞表面蛋白质多糖的分布变化[13]。
1.3.3 DFT计算探究驯化菌在矿物表面的吸附
已有的研究表明,微生物接近并吸附于矿物表面后,更有利于矿物-微生物界面相互作用的发生[14]。因此,微生物对矿物表面的吸附是生物浸出的关键环节。通过DFT模拟计算,探究了驯化菌吸附矿物表面能力的变化。
计算中以黄铁矿为底物,经收敛性测试后,选取截断能Ecut=340 eV,K点为2×2×1,采用超软赝势描述离子实与价电子之间的相互作用,采用GGA-PBE理论方法 [15]。将黄铁矿优化后进行切面,得(100)面,设置15  的真空层,固定底部原子层,驰豫表面后,建立2×2×1超晶胞结构;对于吸附基团,计算时采用 Gamma 点[16];能量收敛标准为2.0×10-5 eV/atom,原子位移收敛标准为0.2 pm,原子间作用力收敛标准为0.8 eV/nm,晶体内应力收敛标准为0.2 GPa;自洽迭代收敛精度为2.0×10-6 eV/atom。所有计算均采用自旋极化方法。
的真空层,固定底部原子层,驰豫表面后,建立2×2×1超晶胞结构;对于吸附基团,计算时采用 Gamma 点[16];能量收敛标准为2.0×10-5 eV/atom,原子位移收敛标准为0.2 pm,原子间作用力收敛标准为0.8 eV/nm,晶体内应力收敛标准为0.2 GPa;自洽迭代收敛精度为2.0×10-6 eV/atom。所有计算均采用自旋极化方法。
2 结果与讨论
2.1 胁迫实验和驯化实验结果
L. ferriphilum菌在不同浓度梯度的As(III)和As(V)下的生长情况如图1(a)和(b)所示。从图中可以看到,未经驯化的L. ferriphilum菌明显受到2.0 g/L As(III)和4.0 g/L As(V)的抑制作用。可以看出,L. ferriphilum菌的耐砷能力并不高,若是不经驯化,难以满足浸出高砷金矿的要求。所以,对L. ferriphilum菌进行驯化是十分必要的。
经驯化实验后,得到能在7.0 g/L As(III)浓度和11.0 g/L As(V)浓度条件下生存的L. ferriphilum菌,所得菌株的耐砷性能均超出了驯化前的菌株,较前人的工作也有所提高[4-6],生长曲线如图1(c)所示。从生长曲线可以看出,未经驯化的L. ferriphilum菌在7.0 g/L As(III) 和11.0 g/L As(V)浓度下基本不能存活;驯化后L. ferriphilum菌在7.0 g/L As(III)条件下,生长至稳定期时,其细胞浓度约为2×108 mL-1,而在11.0 g/L As(V)条件下生长至稳定期的细胞浓度可达到约3×108 mL-1,这表明驯化后L. ferriphilum菌的耐砷能力大大提高;而驯化菌在8.0 g/L As(III)和12.0 g/L As(V)条件下,生长受到了严重的抑制,表明L. ferriphilum菌的耐受能力已经达到了极限。图中驯化菌的细胞浓度在前期有所下降,这可能是驯化菌选择性吸附到矿物表面上[17] ,导致溶液中细胞数量下降。

图1 在不同浓度As(III)和As(V)胁迫下L. ferriphilum菌的生长曲线及驯化前后L. ferriphilum菌的生长曲线
Fig. 1 Evolution of cell concentration of L. ferriphilum under stress of As (III) (a) and As (V) (b) with different concentrations and growth curves (c) of L. ferriphilum before and after acclimation
2.2 As(III)和As(V)驯化菌浸出砷黄铁矿实验效果对比
2.2.1 浸出行为的表征
砷黄铁矿浸出过程浸出液中的细胞浓度、pH、ORP,铁离子和砷离子浓度变化情况分别如图2(a)~(e)所示。由图2可知,As(III)驯化的L. ferriphilum菌浸出液细胞浓度较小,可能是因为As(III)驯化菌吸附到矿物表面上,导致溶液中的细胞浓度较小;pH先增高后降低,氧化还原电位迅速增高,这是由于细菌的生化过程引起的[18],电位在后期下降,这可能是由于溶液pH的变化[19]以及Fe(III)的沉淀等引起的;铁离子和砷离子浓度都是先升高后趋于平稳,数值大小反映了细菌浸出能力的强弱,可以看到,驯化菌对应的数值都大于原始菌的。综合对比图中的数据可以发现,驯化菌的浸出效率总体上高于原始菌的,说明驯化菌更容易适应含砷的环境,驯化达到了预期的目的。浸矿率从大到小的顺序依次为五价砷驯化的菌株、三价砷驯化的菌株、未驯化的菌株。
2.2.2 矿物形貌观察
为更好地对比驯化菌和原始菌浸出砷黄铁矿的能力,根据浸出液测得的数据分析,选取7.0 g/L As(Ⅲ)驯化后的L. ferriphilum菌与原始菌浸出砷黄铁矿过程中的矿渣进行扫描电镜观察,选取无菌体系作为空白对照,其结果如图3所示。
从图3可以看出,36 h之后,驯化菌对砷黄铁矿的腐蚀速度更快,腐蚀程度更大,这说明驯化菌在砷黄铁矿体系中的适应性较原始菌的强,对砷的抵抗力更强,更容易吸附在含砷的矿物上,能更快地浸出砷黄铁矿。
2.3 红外吸收光谱检测
图4所示为各砷浓度梯度驯化后菌的红外光谱图。与文献对比可知[20-23],在2921 cm-1和2853 cm-1处是亚甲基的振动峰,1650 cm-1处是酰胺化合物C=O的特征峰,1560 cm-1处为酰胺键的特征峰,1452 cm-1处是苯环骨架的振动峰。由图4(a)可知,上述位置的峰都随着砷浓度的增加而由尖峰变为钝峰,且峰的强度逐渐减弱,这说明驯化后L. ferriphilum菌的亚甲基减少,驯化过程中C=O键在减少,苯环骨架的量也有所减少,这可能是细菌某些糖类或蛋白质表达量减少所致;1402 cm-1处的峰是糖类C―H的振动峰,850 cm-1处的峰是糖类基团的吸收峰,660 cm-1处的峰是脂肪胺基团的振动峰,这些位置的峰随着砷浓度的增加,由钝峰变为尖峰,强度增大,证明细胞的某些蛋白质、糖类和脂肪胺大量增加;驯化后L. ferriphilum菌480 cm-1处C―C长链的变形振动峰相对较强,也表明经砷驯化后菌株某些多糖或脂质表达量增多。
由图4(a)和(b)可知,从总体上看,随着砷浓度的增加,驯化后浸矿菌的蛋白质,脂质和多糖表达量增加。相对于原始菌,7.0 g/L As(Ⅲ)驯化的L. ferriphilum为As(Ⅲ)的毒性较As(Ⅴ)的毒性要强[24],使菌株的性质变化更大,因此,实验时选取7.0 g/L As(Ⅲ)驯化的L. ferriphilum菌进行驯化机制的研究,能取得更好的结果。

图2 As(III)和As(V)驯化前后的L. ferriphilum菌浸出砷黄铁矿过程中浸出液细胞浓度、pH、氧化还原电位,Fe3+与总铁以及砷离子变化曲线
Fig. 2 Evolution of cell concentration, pH, redox potential, Fe3+, total iron and arsenic ions during bioleaching of arsenopyrite by origin and As(III)/As(V) acclimated L. ferriphilum.

图3 原始L. ferriphilum菌和7 g/L As(Ⅲ)驯化L. ferriphilum菌以及无菌对照体系浸出砷黄铁矿矿渣的SEM像
Fig. 3 SEM images of arsenopyrite slag leached by origin L. ferriphilum ((a1)-(a4)), 7 g/L As(Ⅲ) acclimated L. ferriphilum ((b1)-(b4)) and stile control ((c1), (c2))
2.4 7 g/L As(Ⅲ)驯化后L. ferriphilum菌糖类和蛋白质的含量变化
2.4.1 糖和蛋白质含量的测量
使用BCA法和原位荧光染色法测量驯化菌与原始菌糖类和蛋白质的含量,以去EPS的原生质体为对照,实验结果如图5~6所示。从图5可以看出:较原始菌,驯化菌的糖类和蛋白质的含量都有较大的增加。这与红外光谱(图6)结果一致,也与现有的研究结果相一致[9],表明细菌糖类和蛋白质表达量的增多有利于细菌提高对砷的耐受能力。
2.4.2 7 g/L As(Ⅲ)驯化前后L. ferriphilum菌表面性质的探究
驯化菌表面的电荷量、糖蛋白分布和砷离子的价态变化如图7所示。
图7(a)所示为驯化菌与原始菌的动电位变化情况。随着pH增加,菌株表面的负电荷增多,这是因为细胞表面的羟基发生去质子化,表面正电荷减少,负电荷增加,电位下降[25]。等电点的变化能反应官能团如羟基、羧基和氨基的变化,这些官能团决定着细胞表面的电荷[26]。驯化菌等电点较原始菌等电点低,在pH 1.5附近,这可能是由驯化菌表面带羧基的糖类表达量增多引起的[27],这说明驯化菌表面成分发生了改变;驯化菌电位下降较慢,表明驯化菌表面所带的负电荷较少,更接近中性,这能减弱细胞表面与砷离子间的离子作用力,可能更有利于菌在砷环境下的 生存。
图7(b)所示为驯化菌和原始菌以及它们分别除去EPS后碳的K边XANES谱图,查阅资料得知,特征峰A(288.3 eV)来源于羧基或者氨基1s电子到p*轨道的共振吸收峰[28],特征峰B(292 eV附近)主要来源于σ*cc结构[29]。XANES谱图峰面积的大小与细菌表面糖和蛋白的含量存在正相关关系。由于C元素K边的能量较低[30],穿透能力弱,未去除EPS的驯化菌和原始菌K边XANES主要反应的是胞外EPS中有机组成的信息,而去除了EPS的驯化菌和原始菌K边XANES主要为周质空间的有机组成信息。从图中可知,完整驯化菌的A峰强度较原始菌的强,说明驯化菌EPS上糖类和蛋白质的含量增多;完整未驯化菌较去除EPS的未驯化菌的A峰稍强,B峰强度相近,表明驯化前菌株糖类和蛋白质在EPS和周质空间中分布较为均匀;驯化菌去除EPS前后的A峰显著增强,说明L. ferriphilum菌经驯化后,糖类和蛋白质的分布由均匀分布在EPS和周质空间中变为集中分布在EPS上,说明EPS上糖类和蛋白质的增多,是该细菌抵抗砷的毒害的重要途径。
图7(c)所示为7 g/L As(Ⅲ)驯化后L. ferriphilum菌砷L边XANES 谱图,对比L边XANES 谱图可以确定元素的价态[31]。对比图中曲线可以看到,驯化菌表面的砷主要是以As(Ⅲ)的形式存在,这说明驯化菌的解毒机制可能以甲基外排途径为主[32],也可能是因为细菌未能及时将吸附在表面的As(Ⅲ)转化为As(Ⅴ),导致As(Ⅲ)积累在细胞表面。
2.5 DFT计算探究驯化菌在矿物表面的吸附
利用Materials Studio 软件中 CASTEP 模块的第一性原理对黄铁矿晶胞进行优化后,得到的晶格常数为0.5428 nm,与实验值0.5417 nm很接近[16],只有约0.2%的误差,表明计算结果与实际情况是相符合的。

图4 As(Ⅲ)和As(V)分别驯化的L. ferriphilum菌的红外吸收光谱
Fig. 4 FTIR spectra of L. ferriphilum acclimated by As(Ⅲ) (a) and As(V) (b)
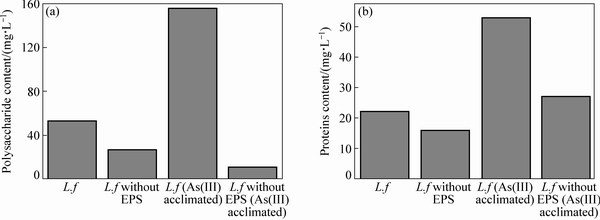
图5 As(Ⅲ)驯化前后的L. ferriphilum菌多糖和蛋白质含量图
Fig. 5 Polysaccharide content of L. ferriphilum before and after acclimation by As(Ⅲ) (a) and protein content of L. ferriphilum before and after domestication by As(Ⅲ) (b)
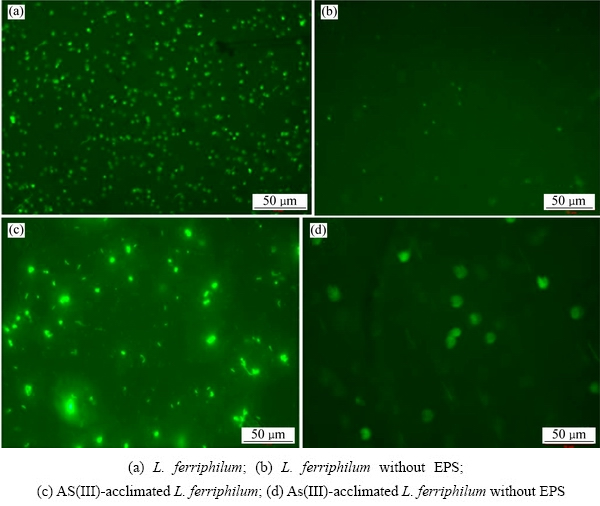
图6 不同处理条件下糖蛋白的荧光梁色图
Fig. 6 Fluorescence staining of glycoprotein by different treatments
由前面的实验得知驯化菌的糖类和蛋白质含量增多,随之增多的是细胞表面的羟基(―OH),以丝氨酸中的羟基为例,利用CASTEP 模块的第一性原理进行羟基在黄铁矿(100)面上的吸附计算,探究羟基对菌株吸附矿物能力的影响,优化后羟基对表面Fe或S的吸附方式如图8所示。
2.5.1 吸附能的计算
根据公式Ead=Esurface+R-OH-Esurface-ER-OH对图7(a),(b)中两种吸附方式的吸附能进行计算,得到R―OH在Fe上吸附时的Ead为-3.63 eV,R―OH在S上吸附时的Ead为-3.57 eV,说明吸附过程在能量上是有利的,即细菌表面―OH容易在黄铁矿表面吸附,并在铁原子上的吸附更容易。
2.5.2 布居分析和分波态密度图(PDOS)分析
表1给出了吸附后的部分布居分析结果。其中键长较小,进一步说明羟基能与黄铁矿(100)面上的铁和硫原子成键;现有研究表明,布居值越大,形成共价键的可能性就越大[33],而表1中 布居值偏向0,说明R―OH在Fe和S上的两种吸附方式(见图8)均偏向于离子键成键作用。

图7 As(Ⅲ)驯化前后L. ferriphilum菌动电位,As(Ⅲ)驯化前后L. ferriphilum菌碳的K边XANES谱图以及驯化菌砷L边的XANES谱图
Fig. 7 Zeta-potential of L. ferriphilum before and after acclimation by As(Ⅲ) (a), C-K-edge XANES of L. ferriphilum before and after acclimation by As(Ⅲ) (b) and As-L-edge XANES of L. ferriphilum acclimated by As(Ⅲ) (c)
本实验中进一步对羟基吸附(100)面后的分波态密度图(PDOS)进行分析,结果如图9所示。其中,EF代表费米面能级。从图9可以看到,吸附后,O 2p轨道与S 2p轨道在-10~0 eV处交叠,而0~5 eV处的峰较弱,说明O―S间存在成键作用;同时,O 2p轨道与Fe 3d 轨道在-8~0 eV处交叠,0~5 eV处的峰较弱,说明存在O―Fe成键作用[34]。这些结果表明羟基以化学吸附的形式吸附在黄铁矿(100)面上,这与布居分析的结果相一致。
表1 部分布居分析结果
Table 1 Parts of Mulliken population analysis


图8 R―OH吸附在Fe和S上的方式
Fig. 8 Adsorbing stratus of R―OH on Fe (a) and S (b)

图9 R―OH吸附S和Fe后的PDOS谱
Fig. 9 PDOS patterns of R―OH adsorbing S (a) and (b)
综上DFT的计算结果,可知羟基通过与黄铁矿之间的成键作用,以化学吸附的方式吸附在矿物的表面上,吸附能力强。因此,驯化菌表面羟基的增多,糖类和蛋白质含量的增多,有利于菌吸附到矿物表面上,这与已有文献的结论相一致[35]。菌株更好地选择性吸附到矿物上,有利于菌更快地获取生长所需能量[36],从而适应高砷的环境,这可能也是微生物抗砷的机制之一。
3 结论
1) 经过驯化,得到了As(III)和As(V)耐受浓度分别为7.0 g/L和11.0 g/L的L. ferriphilum菌,并且驯化菌有良好的砷黄铁矿浸出能力。
2) 通过实验数据可知,驯化菌的糖类和蛋白质表达类型发生了改变,不利于细胞生存的糖类和蛋白质表达量减少,有利于细胞生存的糖类和蛋白质表达量增多,总体上糖类和蛋白质的表达量增多。
3) 动电位表明驯化菌表面的负电荷量减少,通过对碳的K边分析得知驯化菌表面糖类和蛋白质的分布位置发生了变化,从均匀分布转变为主要集中在EPS上,这有利于菌株在高砷环境下生存;通过砷的L边分析得知As在驯化菌表面主要以三价砷存在,由此推测出细菌耐砷机制可能是以甲基外排途径为主,但这个推测还需要进一步的实验进行验证。
4) DFT计算结果表明,驯化菌细胞表面糖类和蛋白质中羟基的增多,有利于菌以化学吸附的方式吸附到矿物上,从而更容易获取能量,从而适应高砷环境。
致谢:本论文的DFT计算工作在吕梁国家超级计算中心天河2号机组以及中南大学和南京大学高性能计算中心完成。
REFERENCES
[1] 杨 硕. 氧化硫硫杆菌耐砷菌株与非耐砷菌株的生理生化特性比较[C]//中国环境科学学会学术年会论文集(第八卷). 北京: 中国环境科学出版社, 2013: 7106-7118.
YANG Suo. Comparison of physiological and biochemical characteristics between arsenic-tolerant strains and non-arsenic-tolerant strains of Thiobacillus ferro- oxidans[C]//Proceedings of the Chinese Society for Environmental Sciences (Volume 8th). Beijing: China Environmental Press, 2013: 7106-7118.
[2] 杨洪英, 范 金, 崔日成, 巩恩普. 难处理高砷金矿的细菌氧化-提金研究[J]. 贵金属, 2009, 30(3): 1-3.
YANG Hong-ying, FAN Jin, CUI Ri-cheng, GONG En-pu. Study on bacterial oxidation-extraction gold of refractory gold ore[J]. Precious Metals, 2009, 30(3): 1-3.
[3] 方 芳, 钟 宏, 江放明, 许凯扬. 嗜酸氧化亚铁硫杆菌的耐砷驯化与浸矿能力[J]. 中南大学学报(自然科学版), 2013(10): 3977-3983.
FANG Fang, ZHONG Hong, JIANG Fang-ming, XU Kai-yang. Domestication for arsenic-tolerant ability and bioleaching of Acidithiobacillus ferrooxidans[J]. Journal of Central South University (Science and Technology), 2013(10): 3977-3983.
[4] ZHAO Qing, LIU Xiang-mei, ZHAN Yang, LIN Jian-qun, YAN Wang-ming. Construction of an engineered Acidithiobacillus caldus with high-efficiency arsenic resistance[J]. Acta Microbiologica Sinica, 2005, 45(5): 675-679.
[5] JIANG Hui-dan, LIANGYi-li, YIN Hua-qun, XIAO Yun-hua, GUO Xue, XU Ying, HU Qi, LIU Hong-wei, LIU Xue-duan. Effects of arsenite resistance on the growth and functional gene expression of Leptospirillum ferriphilum and Acidithiobacillus thiooxidans in pure culture and coculture[J]. Biomed Research International, 2015(18): 1-13.
[6] 吴俊子, 曾伟民, 王玉光, 仉丽娟, 万利利, 周洪波. 搅拌槽反应器中中度嗜热浸矿菌预处理含砷金矿[J]. 过程工程学报, 2013, 13(3): 494-499.
WU Jun-zi, ZENG Wei-min, WANG Yu-guang, LI Li-juan, ZHOU Hong-bo. Pretreatment of arsenic-bearing gold ores in stirred tank reactor by medium thermophilic bioleaching bacteria[J]. The Chinese Journal of Process Engineering, 2013, 13(3): 494-499.
[7] 罗金学. 喜温硫杆菌蛋白质组学研究技术建立及抗砷机制初步探讨[D]. 济南: 山东大学, 2008.
LUO Jing-xue. Construct of proteomic technology for Acidithiobacillus caldus research and study on arsenic resistant mechanlsm[D]. Jinan: Shandong University, 2008.
[8] 冷非凡. 氧化亚铁硫杆菌和氧化硫硫杆菌对无机砷化合物抗性的比较研究[D]. 兰州: 兰州大学, 2010.
LENG Fei-fan. Comparative study on the resistance of Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans to inorganic arsenic compounds[D]. Lanzhou: Lanzhou University, 2010.
[9] 富 瑶. 砷胁迫下浸矿细菌胞外聚合层(EPS)中多糖抗砷行为的研究[D]. 沈阳: 东北大学, 2011.
FU Yao. Study on anti-arsenic behavior of polysaccharide in extracellular polymeric layer (EPS) of leaching bacteria under arsenic stress[D]. Shenyang: Northeastern University, 2011.
[10] LIU Hong-chang, XIA Jin-lan, NIE Zhen-yuan. Relatedness of Cu and Fe speciation to chalcopyrite bioleaching by Acidithiobacillus ferrooxidans[J]. Hydrometallurgy, 2015, 156: 40-46.
[11] ZARNOWIEC P, LECHOWICZ L, CZERWONKA G, KACA W. Fourier transform infrared spectroscopy (FTIR) as a tool for the identification and differentiation of pathogenic bacteria[J]. Current Medicinal Chemistry, 2015, 22(14): 1710.
[12] NIE Zhen-yuan, LIU Hong-chang, XIA Jing-Lan, YANG Yi, ZHEN Xiang-Jun, ZHENG Li-juan, QIU Guan-zhou. Evidence of cell surface iron speciation of acidophilic iron-oxidizing microorganisms in indirect bioleaching process[J]. Biometals, 2016, 29(1): 25-37.
[13] LIU Li-zhu, PAN Xuan, XIA Xu, ZHOU Yu-hang, NIE Zhen-yuan, XIA Jin-lan. In situ characterization of surface organic composition changes of thermoacidophilic archaea Acidianus manzaensis YN-25 in response to energy substrate[J]. Solid State Phenomena, 2017, 262: 417-420.
[14] LIU Jian-she, WANG Zhao-hui, GEN Mei-mei, QIU Guan-zhou. Progress in the study of polyphase interfacial interactions between microorganism and mineral in bio-hydrometallurgy[J]. Mining & Metallurgical Engineering, 2006.
[15] 李 帅, 赵志曼, 郜 峰, 杜晨曦, 刘子瑜. 羧基与钙离子吸附机理的密度泛函的仿真研究[J]. 计算机与应用化学, 2015, 32(8): 951-954.
LI Shuai, ZHAO Zhi-man, GAO Feng, DU Chen-xi, LIU Zi-yu. Simulation study on the adsorption mechanism of carboxyl and calcium ions by density functional analysis[J]. Computer and Applied Chemistry, 2015, 32(8): 951-954.
[16] 李玉琼, 陈建华, 蓝丽红, 郭 进. 氧分子在黄铁矿和方铅矿表面的吸附[J]. 中国有色金属学报, 2012, 22(4): 1184-1194.
LI Yu-qiong, CHEN Jian-hua, LAN Li-hong, GUO Jin. Adsorption of O2 on pyrite and galena surfaces[J]. The Chinese Journal of Nonferrous Metals, 2012, 22(4): 1184-1194.
[17] 王 蕾, 夏金兰, 朱泓睿, 刘红昌, 聂珍媛, 刘李柱. 微生物-矿物相互作用及界面显微分析研究进展[J]. 微生物学通报, 2017, 44(3): 716-725.
WANG Lei, XIA Jin-lan, ZHU Hong-rui, LIU Hong-chang, NIE Zhen-yuan, LIU Li-zhu. Progress on research of microbe-mineral interaction and interfacial microanalysis[J]. Microbiology, 2017, 44(3): 716-725.
[18] 战金帅. 湖南省典型高砷金矿的细菌氧化及提金试验研究[D]. 沈阳: 东北大学, 2005.
ZHAN Jin-shuai. Study on the bacterium oxidations test of the type high arsenic gold concentration from Hunan and the extracted gold[D]. Shenyang: Northeastern University, 2005.
[19] DENG Sha, GU Guo-hua, HE Guo-shuai, LI Li-juan. Catalytic effect of pyrite on the leaching of arsenopyrite in sulfuric acid and acid culture medium[J]. Electrochimica Acta, 2018, 263: 8-16.
[20] 周维芝, 申博玲, 刘升波, 陈 波, 张玉忠. 南极海冰细菌胞外多糖的助凝作用及红外光谱分析[J]. 光谱学与光谱分析, 2009, 29(9): 2405-2408.
ZHOU Wei-zhi, SHEN Bo-ling, LIU Shen-bo, CHRN Bo, ZHANG Yu-zhong. Effect of extracellular polysaccharide of Antarctic sea ice bacteria on coagulation and FTIR analysis[J]. Spectroscopy and Spectral Analysis, 2009, 29(9): 2405-2408.
[21] 赵花荣, 王晓燕, 陈冠华, 温树敏. 利用傅里叶变换红外光谱法鉴定小麦品种[J]. 光谱学与光谱分析, 2004, 24(11): 1338-1341.
ZHAO Hua-rong, WANG Xiao-yan, CHEN Guan-hua, WEN Shu-ming. Identification of wheat varieties by Fourier transform infrared spectroscopy[J]. Spectroscopy and Spectral Analysis, 2004, 24(11): 1338-1341.
[22] 郭振楚, 韩永生, 封惠侠. 三种多糖的光谱鉴定、化学改性及活性[J]. 光谱学与光谱分析, 1999, 19(1): 25-27.
GUO Zhen-chu, HAN Yong-sheng, FENG Hui-xia. Spectral identification, chemical modification and activity of three polysaccharides[J]. Spectroscopy and Spectral Analysis, 1999, 19(1): 25-27.
[23] 周 群, 孙素琴, 杜德国, 梁曦云, 杨显荣. 芦丁热稳定性的红外光谱法实时原位跟踪[J]. 光谱学与光谱分析, 2000, 20(2): 195-198.
ZHOU Qun, SUN Su-qing, DU De-guo, LIANG Xi-yun, YANG Xian-rong. The thermal stability of rutin in situ tracking by infrared spectroscopy[J]. Spectroscopy and Spectral Analysis, 2000, 20(2): 195-198.
[24] 蔡 林, 王革娇. 抗砷性微生物及其抗砷分子机制研究进展[J]. 微生物学通报, 2009, 36(8): 1253-1259.
CAI Lin, WANG Ge-jiao. Advance on studies of arsenic-resistant microorganisms and molecular mechanisms[J]. Microbiology, 2009, 36(8): 1253-1259.
[25] 王 永, 徐仁扣, 王火焰, 姜 军. 砷酸根在可变电荷土壤颗粒表面的配位吸附[J]. 环境化学, 2009, 28(2): 163-167.
WANG Yong, XUN Ren-kou, WANG Huo-yan, JIANG Jun. The coordination adsorption of arsenic on the surface of variable charge soil particles[J]. Environmental Chemistry, 2009, 28(2): 163-167.
[26] XIA Le-xian, LIU Xin-xing, ZENG Jian, CHU Yin, GAO Jian, LIU Jian-she, QIU Guan-zhou. Mechanism of enhanced bioleaching efficiency of Acidithiobacillus ferrooxidans after adaptation with chalcopyrite[J]. Hydrometallurgy, 2008, 92(3): 95-101.
[27] 许晓芳, 林 海, 董颖博, 周闪闪. Mg2+对At.f菌氧化活性的影响[J]. 华南理工大学学报(自然科学版), 2016, 44(6): 41-46.
XU Xiao-fang, LIN Hai, DONG Ying-bo, ZHOU Shan-shan. Effect of Mg2+ on the oxidative activity of At.f bacteria[J]. Journal of South China University of Technology (Natural Science Edition), 2016, 44(6): 41-46.
[28] KEILUWEIT M, BOUGOURE J J, ZEGLIN L H, MYROLD D D, WEBER P K, PETT-RIDGE J, KLEBER M, NICO P S. Nano-scale investigation of the association of microbial nitrogen residues with iron (hydr)oxides in a forest soil O-horizon[J]. Geochimica Et Cosmochimica Acta, 2012, 95(11): 213-226.
[29] ENDO O, NAKAMURA M, AMEMIYA K. Separation of C K-NEXAFS spectra for layer-by-layer analysis of carbon-based thin films: An n-alkane monolayer adsorbed on a monolayer graphene substrate grown on a Pt(111) surface[J]. Journal of Electron Spectroscopy & Related Phenomena, 2013, 189(15): 27-31.
[30] 陈 亮. 同步辐射软X射线辐射效应研究[D]. 合肥: 中国科学技术大学, 2011.
CHEN Liang. Study on soft X-ray radiation effect of synchrotron radiation[D]. Hefei: University of Science and Technology of China, 2011.
[31] LIANG Chang-Li, XIA Jin-lan, YANG Yi, NIE Zhen-yuan, ZHAO Xiao-juan, ZHENG Lei, MA Chen-yan, ZHAO Yi-dong. Characterization of the thermo-reduction process of chalcopyrite at 65 ℃ by cyclic voltammetry and XANES spectroscopy[J]. Hydrometallurgy, 2011, 107(1/2): 13-21.
[32] 李 兵. 嗜铁钩端螺旋菌在砷胁迫条件下的蛋白质组学研究[D]. 济南: 山东大学, 2010.
LI Bing. The proteomics study of Leptosplrlllum ferriphllum under arsenic stress[D]. Ji’nan: Shandong University, 2010.
[33] ZHAO Cui-hua, CHEN Jian-hua, LI Yu-qiong, HUANG De-wei, LI Wei-zhou. DFT study of interactions between calcium hydroxyl ions and pyrite, marcasite, pyrrhotite surfaces[J]. Applied Surface Science, 2015, 355: 577-581.
[34] LI Kui, ZHAO Yao-lin, ZHANG Peng, HE Chao-hui, DENG Jia, DING Shu-jiang, SHI Wei-qun. Combined DFT and XPS investigation of iodine anions adsorption on the sulfur terminated (001) chalcopyrite surface[J]. Applied Surface Science, 2016, 390: 412-421.
[35] 贾春云, 李培军, 魏德洲, 张海荣, 刘 宛. 微生物在矿物表面吸附的研究进展[J]. 微生物学通报, 2010, 37(4): 607-613.
JIA Chun-yun, LI Pei-jun, WEI De-zhou, ZHANG Hai-rong, LIU Wan. Research advances on adsorption of bacteria to mineral surface[J]. Microbiology, 2010, 37(4): 607-613.
[36] 张东晨, 王 涛, 侯志翔, 周倩倩. 矿物表面性质生物调节机理的研究[J]. 洁净煤技术, 2013, 19(1): 5-9.
ZHANG Dong-chen, WANG Tao, HOU Zhi-xiang, ZHOU Qian-qian. Mechanism of microorganism regulating mineral surface properties[J]. Clean Coal Technology, 2013, 19(1): 5-9.
Domestication of Leptospirillum ferriphilum arsenic-tolerant ability and mechanism of domestication
ZHENG Xing-fu1, NIE Zhen-yuan1, XIA Jin-lan1, LIU Li-zhu1, YANG Hong-ying2
(1. Key Laboratory of Biometallurgy of Ministry of Education of China, School of Resources Processing and Bioengineering, Central South University, Changsha 410083, China;
2. School of Metallurgy, Northeastern Univwrsity, Shenyang 110819, China)
Abstract: In order to obtain high arsenic-tolerant ability of Leptospirillum ferriphilum, continuous transfer domestication was adopted, and the acclimation mechanism was studied. The results show that, after domestication, the tolerant abilities to As(Ⅲ) and As(Ⅴ) of L. ferriphilum increase to 7.0 g/L and 11.0 g/L, respectively. The leaching effect of the bacteria acclimated by As(Ⅴ) is significantly higher than that by As(Ⅲ) in the leaching of arsenopyrite. After being acclimated, the contents of polysaccharide and protein of cells significantly increase, while the negative charge on the surface of cells significantly decreases, so as to adapt to the high arsenic environment. The in-situ C-K-edge XANES results show a change in the distribution of polysaccharide and protein. The DFT calculation shows that the adapted bacterial cells are easier to be adsorbed on the mineral surface.
Key words: arsenic; arsenopyrite; pyrite; domestication
Foundation item: Projects(51774342, U1608254) supported by the National Natural Science Foundation of China
Received date: 2018-01-24; Accepted date: 2018-04-20
Corresponding author: NIE Zhen-yuan; Tel: +86-731-88830544; E-mails: zynie@csu.edu.cn,jlxia@csu.edu.cn
(编辑 龙怀中)
基金项目:国家自然科学基金资助项目(51774342,U1608254)
收稿日期:2018-01-24;修订日期:2018-04-20
通信作者:聂珍媛,高级工程师;电话:0731-88830544;E-mail:zynie@csu.edu.cn,jlxia@csu.edu.cn

