
��Ӧ�淨�Ż���ȡ����Ҷ�������ʼ�����ЧӦ
������1, 2������ӿ1, 2������ƽ1, 2��������1, 2�������1, 2��������1, 2
(1. ���ȴ�ѧ ������ѧ�빤��ϵ������ ���ţ�361021��
2. ���ȴ�ѧ ��������Դ�����о��������� ���ţ�361021)
ժ Ҫ��
�����ó�����ȡ�ķ�����ȡ�����廯�����ʣ���ͨ����Ӧ�淨���Ҵ����������������ȡʱ�䡢��Һ�Ⱥ��¶ȵ������Ż�����ͨ��GC/MS��������Ҫ��������Ϊ���ǻ�����������ȷ��������ʡ��õ���ȡ�������ʵ���������Ҵ��������Ϊ72.92%��ʱ��Ϊ160 min��Һ�ϱ�Ϊ40.03:1 mL/g�������¶�Ϊ55 �档�ڴ���������£�����Ҷ�������ʶԵ���С�����������Ϊ91.34%����ģ����ϵ���Ӧֵ93.52%���������ϵõ���ģ�ͽϺõط���ʵ�ʣ�����Ҷ����Ũ��Ϊ0.1~0.4 g/Lʱ�Ե���С���������Ч�����Ż�������������������Ӷ�����
�ؼ��ʣ�
����������С������������������Ӧ�淨��
��ͼ����ţ�Q946.8��TQ459 ���ױ�־�룺A ���±�ţ�1672-7207(2013)09-3940-08
Optimization of extraction and identification of allelochemical from leaves of Salix babylonica by response surface methodology
JIANG Zhongyang1, 2, GUO Peiyong1, 2, CHEN Liping1, 2, XU Wenxiu1, 2, WAN Jinjin1, 2, LI Shixiong1, 2
(1. Department of Environmental Science and Engineering, Huaqiao University, Xiamen 361021, China;
2. Institute of Environmental and Resources Technology, Huaqiao University, Xiamen 361021, China)
Abstract: Ultrasound was used to investigate the effects of ethanol concentration, extraction time, ratio of liquid to solid��and temperature on allelochemical extraction from the leaves of Salix babylonica. The extraction parameters were optimized through the response surface method (RSM). The isolated fractions were characterized by GC/MS, and phenolic acids were identified as the most potential allelochemicals, especially Hydroxy ethyl cinnamate. The results show that the optimized conditions are as follows: The volume fraction of ethanol concentration is 72.92%, the extraction time is 160 min, the ratio of liquid to solid is 40.03:1 mL/g, and the extraction temperature is 55 ��. Under the optimized inhibition, the rate can reach 91.34% in the verification experiment, which is close to the response value (93.52%) with the model fitting. The optimized model by RSM agrees well with the practice. The inhibition of Chlorella pyrenoidosa increases with the increase of phenolic acids mass concentration at 0.1-0.4 g/L.
Key words: Salix babylonica; Chlorella pyrenoidosa; allelochemical; response surface methodology
ˮ�帻Ӫ�����Ӿ絼�����౬���������γ�ˮ������ˮ�����̬���ܺ�����ˮԴ�İ�ȫ��������Ӱ�졣��ˮ����ֱ��Ͷ�ӻ�ѧɱ���(������ͭ)�������е���á�������Ч���Ϻõ����巽��֮һ������ɱ������ܶ�ʱ��ﵽɱ�����Ч�����������γɶ�����Ⱦ�����⣬����Σ����̬����������Σ�����ཡ��[1]������ֲ������Ļ�����������������������и�Ч����̬��ȫ�ԽϺõ��ص㣬ΪѰ�һ����Ѻõ����巽���ṩ��һ��˼·[2-3]���������ü�һ��ֲ��ͨ�������ͷŻ�ѧ����(��������)Ӱ��������������������[4]��Ŀǰ������ֲ�ﻯ�����ü����������������������ķ�ʽ��Ҫ����3�ࣺ(1) ��ˮ������ֲֲ�ʹ����ˮ���з��ڻ��������������������������˹���������[5]��(2) �������ֲ����Ͷ��������ˮ���У�ֲ���帯�ܽ�����ͷŻ����������壬�����ո�[6-7]��(3) ��ֲ��������ȡ��������Ͷ�ӵ�������ˮ���н����������[8-9]�����У����һ�ʽ������㣬��ҵ�������������Ƴ�[4, 10]����Ŀǰ�÷�����о���Ҫ�����ڳ�ˮֲ��[11-13]��ͦˮֲ��[14-16]��ˮֲ��[17]��ˮ��ֲ�����½��ֲ�������Ǵ���ľ��ֲ����о����٣���������Ӧ�淨��������������ȡֲ���������ʵ��о����١��ڴˣ����������Թ㷺�ֲ��ںӰ��ԡ��������������������Ϊ�о��������ø÷����������ȡֲ���������ʣ���������Ҫ�ɷ֣��о��������ʶԵ���С����������Ӱ�죬̽�ֿ��ܵ��������������Ϊ����½��ֲ�ﻯ�����ʿ���ˮ���ṩ�ο���
1 ʵ����Ϻͷ���
1.1 ʵ����ϼ��Լ�
����ҶƬ���Ի��ȴ�ѧУ���̻�����ȡ�²ɼ�������ҶƬ������60 ������и���48 h���÷�������飬���档����С����(Chlorella pyrenoidosa)�����й���ѧԺ�人ˮ������������SE������[18]�С�����ǰ����ֱ�����������õ�SE��������ʹ֮��ָ�������ں��������顣
�����е��Լ���Ϊ�������������������ܼ��״�Ϊɫ�״���
1.2 ����Ҷ�������ʵ���ȡ��������������
��ȡ����Ҷ��ĩ�ھ�����ƿ�У������Ҵ���Һ����������ȡ(��������Ϊ250 W)��������������ȡҺ�Ⱥ���ֽ���ˡ���Ϊ0.22 ��m�л�ϵ��Ĥ��������ȥ��������õ���Һ��Ȼ��ͨ����ת������ȥ�ܼ����õ����࣬���á�
�������̣�����Ҷ��ĩ���Ҵ�������ȡ������Ũ��������������ȡ���轺������(�����ȼ���ͼ״�Ϊ��ϴ�������Ϊ3:1)���״��ؽᾧ����������(�ṹ����)��
1.3 ��ȡ�����Ż�����
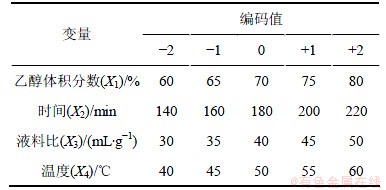
ǰ�ڵ��������鷢�֣����Ҵ��������Ϊ70%��ʱ��Ϊ180 min���¶�Ϊ50 ���Һ�ϱ�Ϊ40.03:1 mL/gʱ������Ҷ���������нϺõ��������á���ˣ���ʵ��������4��������Ϊ���������Design Expert 7.0ͳ�Ʒ���������������������(central composite design (CCD))Ϊ��������Ӧ����������ʵ�飬�Ի�ȡ����ʵ����������������ˮƽ���������1��
��1 ������������������ˮƽ��
Table 1 Range of different factors investigated with CCD design
1.4 ������ֵ�����ɫ��-����(GC-MS)�ⶨ
��GC-MS(Agilent 7890A/5975C)�Ի�����ֽ��нṹ������
����������ɫ����HP-5 (�����ھ���Ĥ��Ϊ30 m��0.25 mm��0.25 ��m)������ΪHe���������¶ȶ�Ϊ280 �档������������������Ϊ1 ��L��������Ϊ1 mL/min���������³�ʼ¯��50 �汣��3 min��Ȼ����20 ��/min�ٶ�������170 �棬����2 min�������10 ��/min������280 �棬����3 min��
����(MS)������EI����Դ����������Ϊ70 eV����Ʒ����ֵ��������ݣ�����NIST02���⣬��������ֵĻ�ѧ�ṹ��
1.5 ������Լ����ݴ���
��������Һ���Ը���ȡ��ֽ���������������⡣������������ѹ���0.5 h��Ȼ���װ��250 mL��ƿ�С�ÿƿ��100 mL�����а���5 mL���ڶ��������ڵ�����Һ��0.5 mL����Һ������Һ���� PYX-250Q-B���˹��������ڽ��У������������£��¶�Ϊ25 �棬����Ϊ3 000~4 000 lx���ⰵ��Ϊ14 h:10 h��ÿ��24 hȡҺ1�Σ�����Ѫ���������¼��������ʱ�ڵ�ϸ����Ŀ�����������ʣ��������ʱ�ʾ����������ȡ��RI����ʽ���£�
RI=(1-N/N0)��100%
ʽ�У�RIΪ�����ʣ�NΪ���뻯���������������ܶ�(��/mL)��N0Ϊ���������ܶȡ�
2 ���������
2.1 ��Ӧģ�͵Ľ����������Լ���
��A=(X1-70)/5��B=(X2-180)/20��C=(X3-40)/5��D=(X4-50)/5Ϊ�Ա�����������Ҷ�������ʶԵ���С����(C. pyrenoidosa)��������Ϊ��Ӧֵ(Y)������Ӧ�����ʵ��(X1, X2��X3�������1)������CCD��Ʒ������ý�����2��ʾ����30��ʵ����Ϊ2�ࣺ��1~24Ϊ����ʵ��㣬�Ա���ȡֵΪ�����������ɵ���ά���㣻��25~30Ϊ����ʵ��㼴��㣬Ϊ��������ĵ㡣���ʵ���ظ�6�Σ��Թ���ʵ����
����Design-Expert 7.0�����Ա�2�е����ݽ��ж��λع���ϣ��õ�������������ȡ�ʶ��Ҵ��������(A)��ʱ��(B)��Һ�ϱ�(C)�ͳ����¶�(D)��Ԫ���λع�ģ��Ϊ
Y=76+6.94A-6.02B+10.65C+11.18D+1.69AB+
4.07AC+9.42AD+7.04BC-9.42BD-5.85CD-
12.52A2-3.98B2-8.75C2-9.34D2
��ʽ�У�������76��ʾ����Ҷ�������ʵ�ƽ����������ʵ����Ʒ�Χ�ڴ��ڽϸߵ�ˮƽ��
���������Իع鷽�̿��Ŷȷ������������ϵ��R2=90.56%������90.56%��ʵ�����ݿ��ø�ģ�ͽ��н��ͣ�˵�����̿ɿ��Խϸߡ������Ϊ9.717������4.000��˵��ģ���ź�ǿ����������ָ��ʵ�顣�ɼ����ûع鷽��Ϊ�Ż�������ȡ�������ʵ�ʵ�������ṩ��һ�����õ�ģ�͡�
��ʵ�������з���������������3������ģ��P��0.000 1�������ع�ģ�ͼ�������˵���÷�����ʵ�������Ϻܺã��ܹ���ȷ��ӳ����Ҷ�������ʵ����������Ҵ��������(A)����ȡʱ��(B)��Һ�ϱ�(C)�ͳ����¶�(D)֮��Ĺ�ϵ��B��BC��Y��Ӱ������(P��0.05)��A��C��D��AD��BD��A2��C2��D2��Y��Ӱ�켫����(P��0.01)��AB��AC��CD��B2��Y��Ӱ�첻����(P��0.05)������ʵ�����Ӷ���Ӧֵ���Ǽ����Թ�ϵ���ӱ�3Ҳ��֪�������ض���Ӧֵ��Ӱ��Ӵ���С����Ϊ�����¶�(D)��Һ�ϱ�(C)���Ҵ����������ʱ��(B)��ʧ�����P=0.741 9��0.05����������˵��ģ����ȫ�������������������ϡ�
2.2 ��ȡ���յ���Ӧ��������Ż�
ͼ1(a)~(f)��ʾ��ֱ�۵ط�ӳ�����ض���Ӧֵ��Ӱ�졣��ͼ1���᷽��仯���ܼ��̶ȿ����ж�������֮�佻�����ö���Ӧֵ��Ӱ��̶ȣ�����Ӧ������Խ���ͣ�˵�������ضԻ������ʳ�����ȡ��Ӱ��Խ���������뷽����������һ�¡�
��2 ������ȡ���������������
Table 2��Design and results of tests

��ͼ1(a)���Կ������Ҵ����������ʱ�佻�����ò���������ͬ���Ҵ�����������в�ͬ�ļ��ԡ�������������ԭ����֪����ͬ���Ե���Һ�Ʊػ�Ӱ�쵽�������ʵ���ȡ�ʣ�ͬʱ�����Բ�ͬ����ҺҲ��Ӱ�쳬������Ĺ��̡����Ҵ���������ϵ�ʱ���������ʡ��ǵ�ˮ����������ȡ������������֬���Ի������ʵĽ��������Ҵ��������̫��ʱ�������ܴ����ѹ��֬���������ܳ����ӣ�����ʹֲ��ϸ�������ʺܿ����̣������Ҵ���ֲ��ϸ��������Ӱ���������ȡ�ʷ������͡���������������һ����Χ�ڣ���ȡ�������Ҵ����������������������Ҵ��������Ϊ70%ʱ����ȡЧ����ã������Ҵ������������70%ʱ����ȡ���м�С���ơ�
��3 �ع鷽�̵ķ������
Table 3 Variance analysis of regression equation

��ͼ1(b)���Կ������Ҵ����������Һ�ϱȽ������ò������������Ҵ����������Һ�ϱȵ�������ȡ�ʳ��������Ӻ��С�����ơ�Һ�ϱ�����ȡ�����е�һ����Ҫ���أ����ݴ�������ԭ������Ҫ������Ӱ����������Һ����������������������ƶ�������Һ��ԽС�����������������ԽС���Ӷ������ƶ�����ԽС�����⣬��Һ��Ҳ��Ӱ������Ļ����������Һ�ϱȾ��ܻ��ڽϴ�̶�����ߴ����ƶ��������������ܼ�Ӱ����ȡ��ϵ�Ĵ��Ⱥʹ��ʣ����������ڻ������ʵ���ȡ��ͬʱ������������ɱ����ܼ����ճɱ����Ѷȡ���ˣ�Һ�ϱȲ���̫�ߡ�
��ͼ1(c)���Կ������Ҵ���������ͳ����¶Ƚ������������������Ҵ���������ͳ����¶ȵ�������ȡ�������Ӻ��С�����¶Ƚϵ�ʱ�����ʷ���ƽ���˶�����С����ɢ����С�����¶�����ʱ�����ʷ���ƽ���˶�����Խ����ɢ����Խ�������ܼ������ʵĻ�ϡ����⣬�¶����߶�ֲ����֯�Ľ���Ͷ���Ч�ɷֵ��ܽ�Ҳ�дٽ����ã��ܹ�ʹ���������̣��ƻ�ø�����������ʵĽ��������¶ȹ���ʱ��һ�������ٻ��Գɷֵķֽ⣬ʹ���ܵ��ƻ�������������ԣ������ʵ��ܳ������ӣ������������������㣻��һ����ʹ�Ҵ��ӷ�������ܼ���ʧ���ҽ����Ҵ����������Ӱ����ȡ�ʡ�
��ͼ1(d)���Կ�����ʱ���Һ�ϱȽ���������������ȡ������Һ�ϱȵ����������Һ�ϱȹ��ή����ȡ�ʡ�����ȡ������ʱ����ӳ������͡��ɼ���Һ�ϱȸ���ʱ�����ȡ���нϴ�Ӱ�졣
��ͼ1(e)���Կ�����ʱ��ͳ����¶Ƚ����������������ų��������¶����ӣ�����������ȡ�ʳ������ӵ����ƣ����ų����¶ȵ����ӣ���ȡ�������Ӻ����м�С��������ȡ����ѧ���ۣ�����ʱ�������ڻ��л������ʵij����ɢ�������ҳ�������Һ�ϵ�����Խ��֣�ϸ��������Խ��ȫ��Խ�����ڻ�����������������ʱ����������ʹϸ���ڴ����IJ����P�Һ�ʵȻ�����ȡҺ�У�ʹ��Һ���������࣬�����Ӷ�Ӱ������Ч�ɷֵ��ܳ������⣬����ʱ��ij������ջ����ÿ��ܻ��ƻ���Ч���ԳɷֵĻ�ѧ�ṹ������Ӱ����ȡ�ʡ�

ͼ1 ��ͬ���ض�������Ӱ�����Ӧ��ͼ
Fig.1 Response surface(3D) that effects of different extraction parameters on inhibition rate
��ͼ1(f)���Կ����������¶Ⱥ�Һ�ϱȽ������ò�����������������ȡ���泬���¶Ⱥ�Һ�ϱȵ�����������ӣ����������¶Ⱥ�Һ�ϱȹ��ߣ��ή����ȡ�ʡ�
2.3 ��ѹ���������ȷ������֤����
��ͼ1(a)~(f)��ʾ����Ӧ����ߵ���Կ������������ʶԵ���С�������������һ����Χ�����Ҵ���������������¶ȡ�ʱ�䡢��Һ�ϱȳ�����أ����ǣ����Ҵ��������Ϊ70%~72.5%��ʱ��Ϊ60~180 min��Һ�ϱ�Ϊ42.50:1 mL/g�Լ��¶�Ϊ55 ������ʱ��ȡЧ����á��ɴ˿�֪��ʵ����ѡ�ķ�Χ�ڴ��ڼ�ֵ������Ӧ������ֵ����������ϵĶ��λع鷽����ƫ���ó���ѵ���ȡ�����������£��Ҵ��������Ϊ72.92%��Һ�ϱ�Ϊ40.03:1 mL/g��ʱ����¶ȷֱ�Ϊ160 min��55 �棬�ڴ���������£�����������ȡ����ߣ���Ե���С���������������ֵ�ɴﵽ93.52%��
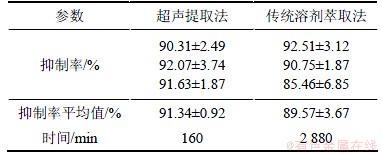
Ϊ����RSM���Ŀɿ��ԣ���������������ȡ������������Ҷ�������ʵ���ȡʵ�顣Ϊ�Աȳ�����ȡ�ͳ�����ᷨ�����ӣ�����ͬһ��ԭ����������ͬʵ�������º�����48 h�õ���ȡҺ����������ԣ��������4���ɱ�4��֪������������ظ�ʵ��3�Σ���������������ƽ��ֵΪ91.34%����Ԥ��ֵ93.52 %�Ͻӽ����ظ��Ժܺã�˵���Ż�����ɿ����봫ͳ��ȡ����ȣ�����������ͬ����³�������ȡʱ��������17�����������ʵ���ȡЧ��Ҳ����һ���̶���ߡ�
��4��������ȡ���볣���ܼ���ȡ����ʵ����
Table 4��Results of ultrasonic and conventional solvent extraction technology
2.4 �������ʽṹ����
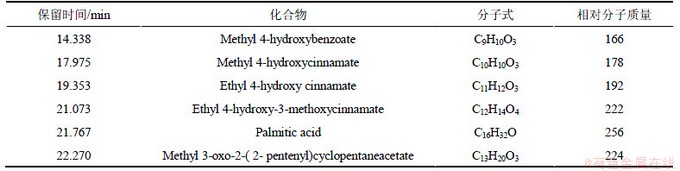
�����봿����Ļ��������ü״��ܽ��0.22 ��m�л���Ĥ���˺�ͨ���������������������5���ӱ�5�ɼ�����Ҫ�������ʰ������������ɸߵ�������Ϊ���ǻ����������(Ethyl 4-hydroxy cinnamate)��4-�ǻ�-3-���������������(Ethyl 4-hydroxy-3- methoxycinnamate)�������(Palmitic acid)�����������(Methyl 3-oxo-2-(2-pentenyl)cyclopentaneacetate)�����ǻ���������(Methyl 4-hydroxycinnamate)�����ǻ����������(Methyl 4-hydroxybenzoate)�������������ֱ�Ϊ12.76%��4.97%��3.35%��2.64%��1.88%��1.07%�����������ǹ���ֲ���ͷŵ�����������ֲ��Ĵ�����л���ʡ�Rice[19]���������ʷ�Ϊ14�࣬Ŀǰ�ձ�Ļ������ʷ�����Ҫ��4��[20-21]��(1) ���ࣻ(2) ���ࣻ(3) �Ǻ������ࣻ(4) �����ͷǵ��װ����ᣬ������Ϊ�������dz���֬���ᡢ�����Լ���������ʡ����о���������Ҷ����������Ҫ���ڷ����������������֬����������(ͳ��Ϊ��������֣���ͬ)��
������,����ֲ��Ļ������������������������о���Ϊ��Ծ[22]�����з����������ǻ��Խ�ǿ��һ�������[23]��Greca��[24]�����������е���������Ϊ����Ͷ�������ʡ�Nakai��[25]���뻨��β���з���������ᡢ�����ᡢ�屶���ᡢ������Ͷ�����ȶ��ַ���������ʣ���֤����ͭ������������н�ǿ���������ã�Wu��[26]�þ���ϩ������ͪPVPP�������塢����Ϳ�ݵķ������еķ������ȥ������3�ֳ�ˮֲ�����ͭ��������������ʷֱ��½�35.7%��43.4%��59.1%��������3�ֳ�ˮֲ���ͷŵķ�������ǽ�����Χˮ������������������Ҫ�������ʡ����о�������Ҷ������η�����ȡ���������õ��Ļ������ʾ�������ҪΪ���������ʣ�����ǻ������������4-�ǻ�-3-����������������ȣ�����Щ��������Ч�����Ƶ���С�����������
2.5 ��������ֶԵ���С����������Ӱ��
ͼ2��ʾΪ��ͬ�����������Ҷ��������ֶԵ���С���������ʱ仯�������ͼ2�ɼ�����ͬ��������ķ�������ֶԵ���С����������������ͬ���������2 d�������ʶ���75%����(P��0.05)�����ֳ����ٸ�Ч������Ч��������Hong��[16]���о���������ƣ�����ڿ���ˮ���廪�������õ���ʵ���塣3 d��0.1~0.3 g/L�����������ʱ����ӳ����в�ͬ�̶��½���0.1 g/L��������������7 d��Ϊ21.2%����0.4 g/L������������ά����89.1~98.2%�����ֳ���ǿ���������á�����������ʱ(7 d)�����������������������������ߣ������˻������ʵ�����Ч�����������������һ���������ԣ����������������Խ��������Ч��Խ�á�
������������һ����з��ǻ��ṹ�Ļ��������廯�����õ���Ҫ���ƿ����Ƿ��������ʵ�������[23, 27]��ʵ��������������һ������Cu2+��Fe3+�ȹ��ɽ������ӣ�����������������ǻ�����������������H2O2[28]��H2O2�ɽ�һ�����ϸ��֬�ʹ�����[29]������ʹ��Ĥ�ṹ��������ϸ����лʧ��������������ͥ͢��[30]�о����������ʶ�ˮ������Ļ�������ʱ���֣���κ��Ͷ��ǻ���������ʹˮ�������弰����С�����ϸ��Ҷ����a�����½���ϸ���ڳ������������ɻ����ߣ�Ĥ֬�ʹ������������ȩ(MDA)���ۣ�ϸ�����������绯ø(SOD)�Ļ�����ʼʱӦ�������ߡ�������[31]�����ڱ����ӺͿ����������ͭ������Ĥ֬�����������������л��������ø���ԣ�ͬʱ����Ĥ�������ԣ�ʹϸ���������ڱ����ӻ�����������ͭ�������SODø���ԣ��ƻ�ϸ������������ϵͳ�����о��е������������������������һЩ���������ʱ����ϵ��ǻ��������������е�Cu2+��Fe3+�ȹ��ɽ������ӽ�ϣ��յ�����������������H2O2������ϸ�����պ��ƻ�ϸ��ø��ϵͳ��Ӱ��ϸ�������������ԣ��Ӷ�������������
��5 ����Ҷ��������GC-MS�������
Table 5 Analytical results of allelochemical from the Leaves of S. babylonica by GC-MS
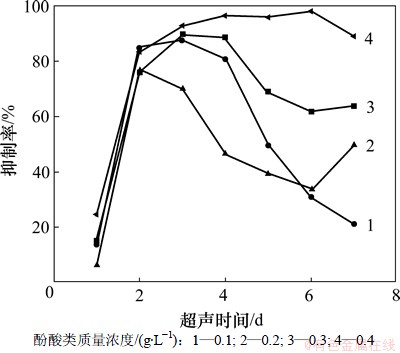
ͼ2 ����������Ũ�ȶԵ���С���������ʵ�Ӱ��
Fig.2 Effects of mass concentration of allelopathic fraction on inhibition rate of C. pyrenoidosa
3 ����
(1) ������ȡ�����������ʵ���ѧ�ع�ģ��Ϊ��
Y=76+6.94A-6.02B+10.65C+11.18D+1.69AB+
4.07AC+9.42AD+7.04BC-9.42BD-5.85CD-
12.52A2-3.98B2-8.75C2-9.34D2
���и������ı���Ϊ��A=(X1-70)/5��B= (X2-180)/20��C=(X3-40)/5��D=(X4-50)/5����������������ȡ��Ӱ��̶ȴӴ���С����Ϊ�����¶�(D)��Һ�ϱ�(C)���Ҵ��������(A)��ʱ��(B)��
(2) ������ȡ�����������ʵ����ʵ���������£��Ҵ��������Ϊ72.92%��Һ�ϱ�Ϊ40.03:1 mL/g���¶�Ϊ55 �棬ʱ��Ϊ160 min��Ԥ��������Ϊ93.52%��ʵ��������Ϊ91.34%��ʵ��ֵ��Ԥ��ֵû���������죬������ѧģ�Ϳ��ԽϺõ�Ԥ���Ҵ����������ʱ�䡢Һ�ϱȺͳ����¶���4�������뵰��С���������ʵĹ�ϵ���ظ��ԽϺá�
(3) ������Ҷ�������ʷ�������õ�����Ҫ�����������Ϊ���ǻ�����������ȷ��������ʣ�������Ũ��Ϊ0.1~0.4 g/L�ķ�������ֶԵ���С����������Ϊ21.2%~89.1%��������������Ͷ�ӻ�����������������������ǿ,���������öԻ������������������һ���������ԡ�
�ο����ף�
[1] Alamsjah M A, Hirao S, Ishibashi F, et al. Isolation and structure determination of algicidal compounds from Ulva fasciata[J]. Bioscience Biotechnology and Biochemistry, 2005, 69(11): 2186-2192.
[2] Ball A S, Williams M, Vincent D, et al. Algal growth control by a barley straw extract[J]. Bioresource Technology, 2001, 77(2): 177-181.
[3] Vyvyan J R. Allelochemicals as leads for new herbicides and agrochemicals[J]. Tetrahedron, 2002, 58(9): 1631-1646.
[4] Rice E L. Allelopathy[M]. 2nd ed. Orlando: Academic Press, 1984: 1-2.
[5] Nakai S, Zou G, Okuda T, et al. Anti-cyanobacterial allelopathic effects of plants used for artificial floating islands[J]. Allelopathy Journal, 2010, 26(1): 113-121.
[6] Newman J R, Barrett P R F. Control of microcystis aeruginosa by decomposing barley straw[J]. Journal of Aquatic Plant Management, 1993, 31: 203-206.
[7] Everall N C, Lees D R. The identification and significance of chemicals released from decomposing barley straw during reservoir algal control[J]. Water Research, 1997, 31(3): 614-620.
[8] Nakai S, Inoue Y, Hosomi M. Algal growth inhibition effects and inducement modes by plant-producing phenols[J]. Water Research, 2001, 35(7): 1855-1859.
[9] Shanab S M M, Shalaby E A, Lightfoot D A, et al. Allelopathic effects of water hyacinth [Eichhornia crassipes][J]. Plos One, 2010, 5(10): 1-8.
[10] ����, ����Ӫ, �ƾ���, ��. ��ͬ�ܼ���ȡ«�����ʶ�ͭ������������Ӱ��[J]. ������ѧ, 2008, 29(11): 3143-3147.
HONG Yu, HU Hongying, HUANG Jingjing, et al. Growth of microcystis aeruginosa affected by allelochemicals of arundo donax Linn extracted with different solvents[J]. Environmental Science, 2008, 29(11): 3143-3147.
[11] Nakai S, Hosomi M. Allelopathic inhibitory effects of polyphenols released by Myriophyllum spicatum on algal growth[J]. Allelopathy Journal, 2002, 10(2): 123-131.
[12] Gross E M. Allelopathy of aquatic autotrophs[J]. Critical Reviews in Plant Sciences, 2003, 22(3/4): 313-339.
[13] Mulderij G, van Donk E, Roelofs J G M. Differential sensitivity of green algae to allelopathic substances from Chara[J]. Hydrobiologia, 2003, 491(1): 261-271.
[14] Li F M, Hu H Y. Isolation and characterization of a novel antialgal allelochemical from Phragmites communis[J]. Applied and Environmental Microbiology, 2005, 71(11): 6545-6553.
[15] He C Q, Wang C K. Allelopathic effect of Acorus tatarinowii upon algae[J]. Journal of Environmental Science, 2001, 13(4): 481-484.
[16] Hong Y, Hu H Y, Sakoda A, et al. Isolation and characterization of antialgal allelochemicals from Arundo donax L[J]. Allelopathy Journal, 2010, 25(2): 357-367.
[17] Sun W H, Yu S W, Yang S Y, et al. Allelochemicals from root exudates of water hyacinth (Eichhornis crassipes)[J]. Acta Phytophysiol Sinica, 1993, 19(1): 92-96.
[18] Bold H C. The morphology of chlamydomonas chlamydogama, Sp. Nov[J]. Bulletin of the Torrey Botanical Club, 1949, 76(2): 101-108.
[19] Rice E L. Allelopathy[M]. New York: Academic Press, 1974: 166-179.
[20] �״���, ����. ֲ�ﻯ��(�������)���ü���Ӧ��[M]. ����: �й�ũҵ������, 2001: 12-42.
KONG Chuihua, HU Fei. Plant allelopathy and its application[M]. Beijing: China Agriculture Press, 2001: 12-42.
[21] �־�, ��ȫ��, �����, ��. ֲ�ﻯ�������о���չ[J]. �й�ũѧͨ��, 2007, 23(1): 68-72.
LIN Juan, YIN Quanyu, YANG Bingzhao, et al. Review on allelopathy of plants[J]. Chinese Agricultural Science Bulletin, 2007, 23(1): 68-72.
[22] Gross E M, Erhard D, Ivanyi E. Allelopathic activity of Ceratophyllum demersum L. and Najas marina ssp intermedia (Wolfgang) Casper[J]. Hydrobiologia, 2003, 506(1/2/3): 583-589.
[23] Inderjit K. Plant phenolics in allelopathy[J]. The Botanical Review, 1996, 62(2): 186-202.
[24] Greca M D, Ferrara M, Fiorentino A, et al. Previtera L. Antialgal compounds from Zantedeschia aethiopica[J]. Phytochemistry, 1998, 49(5): 1299-1304.
[25] Nakai S, Inoue Y, Hosomi M, et al. Myriophyllum spicatum-released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa[J]. Water Research, 2000, 34(11): 3026-3032.
[26] Wu Z B, Gao Y N, Wang J, et al. Allelopathic effects of phenolic compounds present in submerged macrophytes on Microcystis aeruginosa[J]. Allelopathy Journal, 2009, 23(2): 403-410.
[27] Dziga D, Suda M, Bialczyk J, et al. The alteration of Microcystis aeruginosa biomass and dissolved microcystin-LR concentration following exposure to plant-producing phenols[J]. Environmental Toxicology, 2007, 22(4): 341-346.
[28] Tayama S, Nakagawa Y. Cytogenetic effects of propyl gallate in CHO-K1 cells[J]. Mutation Research-Genetic Toxicology and Environmental Mutagenesis, 2001, 498(1/2): 117-127.
[29] Furukawa A, Oikawa S, Murata M, et al. (-)-epigallocatechin gallate causes oxidative damage to isolated and cellular DNA[J]. Biochemical Pharmacology, 2003, 66(9): 1769-1778.
[30] ��ͥ͢, �ⰲƽ, ��÷, ��. ���������ʶ�ˮ������Ļ������ü������[J]. �й�������ѧ, 2007, 27(4): 472-476.
ZHANG Tingting, WU Anping, HE Mei, et al. The allelopathy and its mechanism of phenolic acids on water-bloom algae[J]. China Environmental Science, 2007, 27(4): 472-476.
[31] ����, ������, ����ǿ. �ڱ����ӺͿ������ͭ������Ļ������ü������[J]. ������ѧ, 2008, 27(3): 331-334.
HUA Ming, CHEN Liangyan, YIN Daqiang. The compare on the mechanism of pyrogallic acid and caffeic acid allelochemicals to algae (Microcystis aeruginosa)[J]. Environmental Chemistry, 2008, 27(3): 331-334.
(�༭ �²ӻ�)
�ո����ڣ�2012-08-12�������ڣ�2012-10-23
������Ŀ��������Ȼ��ѧ����������Ŀ(20777021)����������ѧ�����о��ص���Ŀ(210253)������ʡ��Ȼ��ѧ����������Ŀ(2010J01043��D0610012)
ͨ�����ߣ�����ӿ(1969-)���У��ӱ�ʯ��ׯ�ˣ���ʿ�����ڣ�����ˮ��Ⱦ���μ�������Դ�����о����绰��0592-6162300��E-mail: guopeiyong@sina.com
ժҪ��������ҶΪԭ�ϣ����ó�����ȡ�ķ�����ȡ�����廯�����ʣ���ͨ����Ӧ�淨���Ҵ����������������ȡʱ�䡢��Һ�Ⱥ��¶ȵ������Ż�����ͨ��GC/MS��������Ҫ��������Ϊ���ǻ�����������ȷ��������ʡ��õ���ȡ�������ʵ���������Ҵ��������Ϊ72.92%��ʱ��Ϊ160 min��Һ�ϱ�Ϊ40.03:1 mL/g�������¶�Ϊ55 �档�ڴ���������£�����Ҷ�������ʶԵ���С�����������Ϊ91.34%����ģ����ϵ���Ӧֵ93.52%���������ϵõ���ģ�ͽϺõط���ʵ�ʣ�����Ҷ����Ũ��Ϊ0.1~0.4 g/Lʱ�Ե���С���������Ч�����Ż�������������������Ӷ�����
[4] Rice E L. Allelopathy[M]. 2nd ed. Orlando: Academic Press, 1984: 1-2.
[19] Rice E L. Allelopathy[M]. New York: Academic Press, 1974: 166-179.
[20] �״���, ����. ֲ�ﻯ��(�������)���ü���Ӧ��[M]. ����: �й�ũҵ������, 2001: 12-42.
[21] �־�, ��ȫ��, �����, ��. ֲ�ﻯ�������о���չ[J]. �й�ũѧͨ��, 2007, 23(1): 68-72.
[23] Inderjit K. Plant phenolics in allelopathy[J]. The Botanical Review, 1996, 62(2): 186-202.
[30] ��ͥ͢, �ⰲƽ, ��÷, ��. ���������ʶ�ˮ������Ļ������ü������[J]. �й�������ѧ, 2007, 27(4): 472-476.
[31] ����, ������, ����ǿ. �ڱ����ӺͿ������ͭ������Ļ������ü������[J]. ������ѧ, 2008, 27(3): 331-334.
