
���±��: 1004-0609(2006)09-1634-07
�����Leptospirillum ferrooxidans
����Fe2+�ĵ绯ѧ����ѧ
�����1, �Դ�ǿ1, ���庣2, ������2
(1. �����Ƽ���ѧ ұ������̬����ѧԺ, ��������̬��ѭ��ұ���ص�ʵ����, ���� 100083;
2. ������ɫ�����о���Ժ, ���� 100088)
ժ Ҫ��
�о�Leptospirillum ferrooxidans��Һ������ԭ��λ����Fe2+������������ϵ, �õ�������Һln[c(Fe3+)/c(Fe2+)]�����ӳ�����������ϵ; �о�Fe2+�� ϸ��Ũ�ȡ� �¶ȱ仯ʱFe2+������Ӧ���ʵı仯���ɡ� �ڸ���������, ����Һ��Fe2+Ũ�Ƚϵ�ʱ, ����Fe2+Ũ�ȵ�����, Fe2+���������ʼӿ�, ����Fe2+Ũ����5kg/m3����ʱ, Fe2+Ũ�ȵ����Ӳ��������ʹ�������ʼӿ�, ����������Fe2+������; ϸ��Ũ�ȵ�����, ����������֮����, ��ϸ��Ũ����1.25��108cells/dm3����ʱ, ����������ϸ��Ũ�ȵ������нϴ���ȵ�����; ���¶���20~35�淶Χʱ, �¶ȵ�����ӿ�ϸ������Fe2+������, ���¶�������, �������Fe2+�������� ͨ��һϵ�е绯ѧ�����ﻯѧ����, ������Leptospirillum ferrooxidan��������ѧ����, ����������Ea�� Ƶ������K0�� ���۱��ͳ���Km�͵�Ԫ����ϵ��K��i�ȶ���ѧ������ֵ; ����ѧģ���ܺܺ�˵��ʵ������
�ؼ���: Leptospirillum ferrooxidans; ϸ������; ����ұ��; ��ϵ�λ; ����ѧ; �绯ѧ ��ͼ�����: TF803 21; Q939 99
���ױ�ʶ��: A
Electrochemical growth kinetics of Leptospirillum ferrooxidans
through oxidation of Fe2+
LI Hong-xu1, CANG Da-qiang1, DONG Qing-hai2, WANG Dian-zuo2
(1. Ecology and Recycling Metallurgy Laboratory of National Education Ministry,
School of Metallurgy and Ecology Engineering, University of Science and Technology Beijing,
Beijing 100083, China;
2. General Research Institute of Nonferrous Metals, Beijing 100083, China)
Abstract: The relatively relationship between solution rexdox potential and the ferrous iron oxidation rate was established, and it shows that the �� value increases linearly with ln[c(Fe3+)/c(Fe2+)]. All the ferrous ion, the bacteria cells concentration and temperature have apparent effect on the ferrous oxidation rate. When the concentration of ferrous is less than 5kg/m3 the oxidation of ferrous ion increases, however the increase of ferrous ion concentration up to 5kg/m3 would give inhibition of oxidation rate, as well as at the temperature range of 20�� to 35��, the increased temperature can enhance the ferrous ion oxidation, but when the temperature is above 40��, the growth of bacteria inhibis and the oxidation decreases relatively. The increase of cells concentration of bacteria enhances the oxidation apparently especially when the concentration of cells increases to above 1.25��108cells/dm3. Based on the ferrous ion oxidation and redox potential the growth kinetics models were established and the value of activation energy (Ea), frequency factor (K0), saturation constant (Km), and substrate inhibition coefficient (Ki) were determined. The kinetics model is well in coincidence to the experiments results.
Key words: Leptospirillum ferrooxidans; bioleaching; biohydrometallurgy; redox mixed potential; kinetics; electrochemistry
����ұ����һ�Ž�����ѧ�� ұ��ѧ��ѧ�ƽ�������Ŀ�����Դ��̬����ȡ�¼���[1], ��ʵ�ʵ���ѽ��������������, �����õ���Acidthiobacillus ferrooxidans�� Leptospirillum ferrooxidans�ȶ��־���[2, 3]�� �����Խ�������о���Ҫ������Acidthiobacillus ferrooxidans��, �������������ֵ��о���Խ���[4-7]; ��Acidthiobacillus ferrooxidans������Fe2+�� ������Ԫ���������������ͬ, Leptospirillum ferrooxidans��ר��������Fe2+����ɴ�л�Ľ������[8]�� ϸ������Fe2+���зdz���Ҫ������: һ����, ��Ϊ��ԴFe2+��������Ϊϸ���������ṩ�㹻������; ��һ����, Fe2+���������������Fe3+, Fe3+����Ϊ����������������, ������������Fe2+�ֻᱻϸ������ΪFe3+�Թ�����, ��˷���, ��һ�����ֳ�ϸ������ļ�����á� ����ϸ��������, ������ϵ��ϸ��������������, Fe2+������������ͬ, ������ϵ��������λҲ����֮�����ϴ�仯, ���Ȼ����Ľ��������ϴ�Ӱ��[9-11]�� һ����Ϊ, ��Һ��λԽ��, Խ����������������ֽ�, ����Һ��λ��c(Fe3+)/c(Fe2+)�仯��ֱ��Ӱ��, ͬʱ��ϸ���������¶ȵȻ������[11-14]�� ���о������ڽ���ϸ���������к���, ����Ҫ���õIJ���Acidthiobacillus ferrooxidans, ����Leptospirillum ferrooxidans[15, 16]�� ���ܹ���Leptospirillum ferrooxidans����������Խ����Ӱ���о���Щ����[20, 21], ����ϸ����������Һ��λ�仯�Ĺ�ϵ�� ��������Ũ�ȡ� ϸ��Ũ�ȡ� �¶����������ʹ�ϵ����, �����о�Fe2+������ϸ�������ĵ绯ѧ����ѧ��ϵ�IJ�������� �������߽�������Һ��Fe3+/ Fe2+��Ե�λ�ı仯, ��ϵͳ���о�Leptospirillum ferrooxidans��������Fe2+�����Ķ���ѧ��
1 ʵ��
�������Leptospirillum ferrooxidans�����Կ��ˮ�з��롢 �����õ�, ������ʵ����Ҫѱ���� Leptospirillum ferrooxidans���������ñ�9K������, �����Ϊ: 3.0g/L (NH4)2SO4; 0.1g/L KCL; 0.5g/L K2HPO4; 0.5g/L MgSO4��7H2O; 0.01g/L Ca(NO2)3; 9g/L Fe2+; �����Լ���Ϊ������, ��������ȥ����ˮ����, ������pH������Ҫ��6mol/L H2SO4���ڡ�
ϸ���������ö�̬������ʽ�� ��ת�����ξ��нϸ��ԵĴ�Leptospirillum ferrooxidans�������500mL����200mL 9K��������ҡƿ��, ����ҡ���ж�̬����, ����ҺpHά����2, �����ڼ䱣���¶Ⱥ㶨, ҡƿת��200r/min, �������������ܸ�����Һ����, ��֤���Ĺ�Ӧ, ʹ�䲻Ϊϸ���������̶���ѧ�������ء�
��Һ��ϸ�������������������ⶨ, Fe2+���õζ������, ��Һ��������ͨ��ԭ�����շ��ⶨ�� ��Һ��ϵ�λ����ʱ, ����Ƭ�缫��αȵ缫(Ag/AgCl)����ϸ������Һ��, ����ʹ�αȵ缫������Ƭ�缫, ��Һ��ϵ�λͨ��pH��λ�Ʋ�����
2 ���������
Leptospirillum ferrooxidans��ר��������Fe2+����ɴ�л�Ľ������, Fe2+������Ӧ����������ϸ����ĵ绯ѧ��Ӧʽ�����, ����̿�������3ʽ��ʾ

�������������ض�ֻ���������ӵ�������ԭ��ϸ������Һ�л�ϵ�λ��Ӱ��, ����Һ��ϵ�λ������ʽ��ʾ

ͼ1��ʾΪ�ڲ�ͬ�¶���, ������ͬ��c(Fe3+)��c(Fe2+), �������õ�����Һ��λ��ln[c(Fe3+)/c(Fe2+)]�Ĺ�ϵ�� ��ͼ��֪, �ڲ�ͬ���¶���, ���߳����Թ�ϵ, ��ʽ(4)������ �Ҵ�ֱ�ߵ�б�ʿɵæ�0��ֵ, ��ԼΪ695mV, ���ܵ���������ԭ��Fe3+/Fe2+�ı�����ֵ771mV, ����Pesic��[19]��ʵ��ֵ(687mV)�ӽ��� ��ʽ(4)��֪, ����ϸ��������, Fe2+��������Fe3+, ϸ��������֮����, ����Һ�ĵ�λ��֮���������� ��һ���¶���, ������ԭ���Fe3+/Fe2+�Ħ�0ֵ��һ����, �����æ�ֵ, ����Һ�е�����Ũ����֪, �Ҳ�����ϸ���������ڻƼ��������ɵ�Ӱ��ʱ, ���֪ϸ����������������ҺFe2+����Ũ�ȵı仯״���� ��Ϊ�ڲ�ͬ������ѧ�����½�һ���о�ϸ��������������Fe2+�Ķ���ѧ�ṩ�˿��ܡ�

ͼ1 ��ϵ�λ��ln[c(Fe3+)/c(Fe2+)]�Ĺ�ϵ
Fig.1 Relationship between mixed potential and ln[c(Fe3+)/c(Fe2+)
��������Leptospirillum ferrooxidans��ʱ, ��Һ�ĵ�λ�������ֲ���, ���ڳ�����, Fe2+�Ļ�ѧ�����Ǻ����ġ� ��������ϸ����, �ڲ�ͬϸ��Ũ������Һ��λ��ͬ�̶ȳ��������ơ� ͼ2��ʾ�Dz�ͬ��ʼFe2+Ũ��, ������ϸ��Ũ����4.5��108cells/dm3ʱ, ��Һ��ϵ�λ��ʱ��Ĺ�ϵ, ��ʱ�����¶�Ϊ35�档 ��ͼ��֪, ����ϸ����������ֳ, Fe2+������ΪFe3+, Fe3+/Fe2+��λ������ �ڽϸߵij�ʼFe2+Ũ��ʱ, �����ڸ�����Һ��ϸ�����нϿ�������ٶ�, ��ʱ�н϶��Fe2+������, ��c(Fe3+)/c(Fe2+)����Ȼ����, �ʳ�ʼFe2+Ũ����30.0kg/m3ʱ����Һ�ĵ�λ�ϳ�ʼŨ��0.25kg/m3ʱС�� �ɼ���Һ�ĵ�λ��ʱ��ı仯������ֱ����������Fe2+��ϸ���������ʼ�ϸ����������ѧ�Ĺ���, Ҫ���������, ���һ���о�Fe2+������������ҺFe2+Ũ�ȵĹ�ϵ��
ͼ3��ʾΪ�¶Ⱥ㶨Ϊ35��ʱ, ��ͬϸ��Ũ����Fe2+������������ҺFe2+Ũ�ȵĹ�ϵ�� ��ͼ

ͼ2 ��ͬ��ʼFe2+Ũ��ʱ��Һ��λ��ʱ��ı仯
Fig.2 Variation of mixed potential of culture solution of different initial Fe2+ concentration with growth time

ͼ3 �¶�35�桢 ��ͬϸ��Ũ����Fe2+����������
��ҺFe2+Ũ�ȵĹ�ϵ
Fig.3 Relationship between Fe2+ oxidation rate and its concentration under different bacteria concentration at 35��
�п��Կ���, ����ϸ��Ũ�ȵ�����, Fe2+���������ʼӿ�, ������ͬ��Fe2+��, ϸ��Ũ��Ϊ4.50��108cells/mLʱ, Fe2+�������������Կ���ϸ��Ũ��Ϊ0.35��108cells/mLʱ�� ͬʱ, ��ÿһϸ��Ũ����, ����������Fe2+Ũ�ȱ仯��һ��С��������С�Ĺ���, ��������һ��ֵ������������ʵĹ���, ���ͼ2�����ͬ, ˵��Fe2+��ʱ, ��Һ��λ��һ�����
��һ����������¶���ѧ��Ӧ���̽��͡� ����ÿһϸ������һ������ø(E)��������Fe2+, �ɹ�ϸ��������Fe2+Ũ��ΪS, ���ɵ�Fe3+Ũ��ΪP, ����Ϊϸ�������Ļ���Fe2+��Ũ�Ⱥ�ϸ���������ʵĹ�ϵ�������¼��������ʾ:
ϸ������ø��Fe2+���
![]()
��Ϻ�Fe2+������ΪFe3+
![]()
��Ӧ�м���ES��S���ɷǷ�Ӧ�м���ES2
![]()
��������Ӧ���̿��Կ���, ��ϸ������ø��Fe2+�Ӵ�ʱ, ����øͨ����Ӧ(5)�������ӽ��, ��ʱ, ����ø�����ܵ���Fe2+���, ����ͨ����Ӧ(6)��Fe2+������Fe3+ͬʱ�ͷų�������ø, ������ø�ٽ��Fe2+����, ��һ������������Ӧ, ��˷���; ��Fe2+Ũ�Ƚ�Сʱ, Fe2+Ũ�ȵ����ӻ�ӿ췴Ӧ(5)��(6)�Ľ���, ����ʱFe2+Ũ�ȵ����ӻ�ٽ�Fe2+��������Ӧ; ����Fe2+�����Ӻͷ�Ӧ�Ľ���, ��Ӧ(5)��(6)��ﵽһ����̬ƽ��, ��ʱ�練Ӧʽ(3), ����ķ�Ӧ����S���ES��һ�����ΪES2, ��ʱ, Fe2+�����Ӳ�������ٽ�Fe2+������, �����ή����������, ֱ����Ӧ(7)�ﵽ��̬ƽ�⡣ �����ͼ2��3�б��ֳ���Fe2+Ũ������, �������������λ�����½���ԭ��
��Fe2+�������ʿ���������Michalis-Menten���̱�ʾΪ[4, 5]

��Ӧ�ﵽƽ��������dz�ʼ����ø�ͻ���Fe2+Ũ�ȵı�ֵE0/S0�㹻С, ������Fe2+Ũ�Ⱥ�����øŨ����Ȳ��㹻��ʱ, ��Ӧ�ﲻ��ƽ��, ���������費������ ����ϸ����������Fe2+������������������Ҳ�в�ͬ����, Kelly��Jones�Խ����Thiobacillus ferrooxidans�������о������Ϊ, ��Fe2+Ũ����5.6kg/m3����ʱ, ��Ũ�ȵ�������������������, �����ڸ�Ũ��, Ũ�ȵ��������ϸ������������������[5]��
ϸ��Ũ�ȶ�Fe2+�������ʵ�Ӱ����ͼ3���ѵõ���Ӧ�� ��ͼ3��֪, ��һ�����¶���, ����ϸ��Ũ�ȵ�����, ͬFe2+Ũ��ʱ��Fe2+����������֮����, ��ϸ��Ũ����0.35��108cells/dm3���ӵ�4.50��108cells/dm3ʱ, Fe2+�������������(�������ʷ�ֵ)��0.65kg/(m3��h)��ߵ�5.85kg/(m3��h), �ҵ�ϸ��Ũ�����ӵ�1.25��108cells/dm3����ʱ, ����������ϸ��Ũ�ȵ������нϴ���ȵ������� �ɼ�, ҪʹFe2+�������������, ��Ҫ��֤��Խϸߵ�ϸ��Ũ�ȡ�
���ݷ���(8)��ʵ�����ݽ��д���, ����ͼ3�е����ݽ��з����Իع�, ���Եõ�ϸ�������Ķ���ѧϵ��, ���1��ʾ��
��1 ��ͬϸ��Ũ����ϸ������Fe2+�Ķ���ѧ���ʳ���
Table 1 Kinetics rate constant of bacteria oxidizing Fe2+ under different bacteria concentrations

�ɱ�1���ݿ�֪, ϸ��Ũ�ȶԷ�Ӧ���ʳ���K��Ӱ�첻��, ������1.60��108~1.65��108kg��m-3��h-1��cells-1��dm3֮��, �����㶨; ���Ա��ͳ���Km�ͻ�������ϵ��KiӰ��ϴ�, ��ϸ��Ũ������ʱ, Km��Ki��ֵ����������, ����Ki��������Ϊ���ԡ� ��ϸ��Ũ����0.35��108cells/dm3���ӵ�4.50��108cells/dm3ʱ, Km��0.31kg/m3���ӵ�1.21kg/m3, Ki��25.6kg/m3���ӵ�70.4kg/m3, ˵��ϸ����Ũ�ȵ�������Ӱ��������������Ӧ��ƽ�⡣
ϸ��Ũ����Km�仯��ϵ��ͼ4��ʾ�� ��ͼ��֪Km��ϸ��Ũ�ȵı仯������������ϵ�� ����![]() , ��ɽ�ʽ(8)�任Ϊ
, ��ɽ�ʽ(8)�任Ϊ

�÷�Ӧ���ʷ����ܽϺ÷���ͼ4�е�ֱ��, �ɽϺ�˵��ϸ������Fe2+�ķ�Ӧ����ѧ����, ��ֱ�ߵ�б�ʺͽؾ�ɵõ�������ѧ����K��m��K��i�Ľ���ֵ�ֱ�Ϊ0.07kg/m3��2.70cells/dm3��

ͼ4 ϸ��Ũ���뱥��ϵ��Km�Ĺ�ϵ
Fig.4 Relationship between bacteria concentration and saturation coefficient
�������ݿ�֪, ����ϸ��Ũ�ȵ�����, Ki��֮����, ������Ϊ, ϸ��Ũ�ȵ����ӻή�ͻ��ʵ�����ЧӦ, ��ϸ��Ũ������������ϸ������������ø��������, ���ڽϸߵ�Fe2+Ũ���´ﵽ��̬ƽ�⡣ ��Ki�ĵ�����ϸ��Ũ�ȱ仯��ͼ, ��ͼ5��ʾ, 1/Ki��ϸ��Ũ�ȵĹ�ϵ�ɱ�ʾΪ

ͼ5 ϸ��Ũ����1/Ki�Ĺ�ϵ
Fig.5 Relationship between bacteria concentration and K-1i
![]()
����ͼ��ֱ�ߵ�б�ʺͽؾ�, �������ֵΪ27.1kg/m3, ��ֵΪ8.0��108cells/dm3��
��ͬ�¶���Fe2+������������Ũ�ȵĹ�ϵ��ͼ6��ʾ�� ��ͼ�����߿�֪, ���¶���20~35�淶Χʱ, �¶ȵ�����ӿ�ϸ������Fe2+������, ���¶�������, �������Fe2+������, ���¶ȴﵽ40��ʱ, ������������λ��35��30��������������֮�¡� ͬ��ͼ��������ʾ��Fe2+������ЧӦ, ��ʼ����Fe2+Ũ�ȵ�����, �������ʼӿ�, ���ﵽ���ֵ��, ������������Fe2+Ũ�ȵ������������½�����, ��ͬǰ����һ�¡�

ͼ6 �¶ȶ�ϸ������Fe2+���ʵ�Ӱ��
Fig.6 Influence of temperature on oxidation rate of Fe2+ by bacteria
ͨ��ʵ�����ݺ�Ӧ�÷���ʽ(9)����ȷ����ͬ�¶������ʳ���K��ֵ�� ��Ӧ����ѧ�¶ȵ�����õ��ϸߵ�Kֵ�� ���ݻ�ѧ��ø��Ӧ��Arrhenius����, ��Ӧ���ʳ���K���¶ȵĹ�ϵΪ
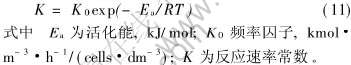
ȡ����, ��lnK��1/T��ͼ, ��һֱ����ͼ7��ʾ�� ����ֱ�ߵ�б�ʺͽؾ�, �õ����EaΪ73.5kJ/mol, Ƶ������K0ֵΪ6492kmol��m-3��h-1��cells-1��dm3�� Ӧ��ͬ���ķ���, Ahonen��[20]�о���T.ferrooxidans�����Ļ��, ��ֵһ����33��96kJ/mol, ���ζ�Leptospirillum ferrooxidans��ܵĽ�����䷶Χ��, �ɹ��ο���

ͼ7 lnK��1/T�Ĺ�ϵ
Fig.7 Relationship between lnK and T-1
��ʽ(10)�� (11)����ʽ(9)�ɵ�ϸ������Fe2+�������ı任��ʽ�Ķ���ѧ����ʽ:

ʽ�е�K0�� Ea�� K��m�� K��i�� ���� �µȶ���ѧ������ֵ����ǰ��ֱ�õ�, ����ʽ(12)��õ���������Leptospirillum ferrooxidans����Fe2+�������Ķ���ѧ����, ���ܺܺõ�˵��ʵ������
3 ����
1) ��Һ������ԭ��λ��������Leptospirillum ferrooxidan��������Fe2+���������������ڵ���ϵ, �о���Ϊ��Һ��ϵ�λ������Һln[c(Fe3+)/c(Fe2+)]�ı仯������������ϵ��
2) ����Һ�¶Ⱥ㶨ʱ, һ����ϸ��Ũ����, ��Һ��Fe2+Ũ�Ƚϵ�ʱ, ���¶�35��, c(Fe2+)��5kg/m3����ʱ, ����Fe2+Ũ�ȵ�����, Fe2+���������ʼӿ�, ����c(Fe2+)��5kg/m3����ʱ, ����Fe2+Ũ�ȵ�����, Fe2+�����������½�, ������һ�����������ֵ��, Fe2+Ũ�ȵ����Ӳ��������ʹ�������ʼӿ�, ������Fe2+���������������á� ����������Һ�е�Fe2+��ϸ�������������ø��Ϲ��̴��ھ���������ϵ, ���ﵽFe2+��������ø�Ľ�ϼ�Fe2+�������� ���ɵ�Fe3+�Ѹ��Ķ�̬ƽ���, Fe2+�����ӻ�����������á�
3) ��һ�����¶���, ����ϸ��Ũ�ȵ�����, ����������֮����, ��ϸ��Ũ����0.35��108cells/dm3���ӵ�4.50��108cells/dm3ʱ, Fe2+�������������(�������ʷ�ֵ)��0.65��ߵ�5.85kg��m-3��h-1, �ҵ�ϸ��Ũ�����ӵ�1.25��108cells/dm3����ʱ, ����������ϸ��Ũ�ȵ������нϴ���ȵ������� ���¶���20~35�淶Χʱ, �¶ȵ�����ٽ�ϸ������Fe2+������, ���¶��ٴ�, �������Fe2+������, ���¶ȴﵽ40��ʱ, ��������������35��30��������������֮�¡�
4) ͨ��һϵ�е绯ѧ�����ﻯѧ����, ������Leptospirillum ferrooxidan��������ѧ����, ������˶���ѧģ���л�ܡ� Ƶ�����ӵȸ�����ֵ��
REFERENCES
[1]Fernando Acevedo. Present and future of bioleaching in developing countries[J]. EJB Electronic Journal of Biotechnology ISSN, 2002, 5(2): 196-199.
[2]�����, ������. ����ұ���е����P������[J]. ��ɫ����, 2003, 55(2): 60-63.
LI Hong-xu, WANG Dian-zuo. Review of investigation on microoganism behaviors in ore bioleaching[J]. Nonferrous Metals, 2003, 55(2): 60-63.
[3]Suzuki I. Microbial leaching of metals from sulfide minerals[J]. Biotechnology Advances, 2001, 19: 119-132.
[4]Lang D S, Lawson F. Kinetics of the liquid-phase oxidation of acid ferrous sulfate by the bacterium Thiobacillus ferrooxidans[J]. Biotechnol Bioeng, 1970, 12: 29-50.
[5]Eccleston M, Kelly D P. Oxidation kinetics and chemostat growth kinetics of Thiobacillus ferrooxidans on tetrathionate and thiosulfate[J]. J Bacteriol, 1978, 134: 718-727.
[6]Dispirito A A, Tuovine O H. Kineticies of uranous and ferrous iron oxidation by Thiobacillus ferrooxidans[J]. Arch Microbiol, 1982, 133: 33-37.
[7]Nagpal S. A structured model for Thiobacillus ferrooxidans growth on ferrous iron[J]. Biotechnology and Bioengineering, 1997, 53: 310-319.
[8]Mignone C F, Donati E R. ATP requirements for growth and maintenance of iron-oxidizing bacteria[J]. Journal Biochemical Engineering, 2004, 18: 211-216.
[9]�����, ������, �¾���. ϸ������ļ�����÷���[J]. ��ɫ����, 2003, 55(4): 98- 100.
LI Hong-xu, WANG Dian-zuo, CHEN Jing-he. Discussion on indirect mechanism of bioleaching[J]. Nonferrous Metals, 2003, 55(4): 98-100.
[10]�����, ������, �¾���, ��. ϸ���������÷���[J]. ��ɫ����, 2003, 55(3): 68-71.
LI Hong-xu, WANG Dian-zuo, CHEN Jing-he, et al. Description of microbe effects on sulfide mineral bioleaching[J]. Nonferrous Metals, 2003, 55(3): 68-71.
[11]�����, ������. ��ϸ���������̵ĵ绯ѧ(��)[J]. ��ұ, 2003, 12(2): 45-48.
LI Hong-xu, WANG Dian-zuo. Electrochemistry of sulfide bioleaching(��)[J]. Mining and Metallurgy, 2003, 12(2): 45-48.
[12]�����, ������. ��ϸ���������̵ĵ绯ѧ(��)[J]. ��ұ, 2003, 12(1): 49-51.
LI Hong-xu, WANG Dian-zuo. Electrochemistry of sulfide bioleaching(��)[J]. Mining and Metallurgy, 2003, 12(1): 49-51.
[13]LI Hong-xu, WANG Dian-zuo, QIU Guan-zhou, et al. Growth kinetics of Thiobacillus ferrooxidans in bioelectrochemical cell[J]. J Cent Technol, 2004, 11(1): 36-40.
[14]�����, ������, ������, ��. Fe2+��T.f�����η�ĩ�缫���������ĵ绯ѧ[J]. �й���ɫ����ѧ��, 2002, 12(6): 1263-1267.
LI Hong-xu, WANG Dian-zuo, HU Yue-hua, et al. Electrochemistry on oxidation of Fe2+ on Thiobacillus ferrooxidans modified powder microelectrode[J]. The Chinese Journal of Nonferrous Metals, 2002, 12(6): 1263-1267.
[15]Falco L, Pogliani C, Curutchet G, et al. A comparison of bioleaching of covellite using pure cultures of Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans or a mixed culture of Leptospirillum ferrooxidans and Acidithiobacillus thiooxidans[J]. Hydrometallurgy, 2003, 71: 31-36.
[16]Boon M, Brasser H J, Hansford G S, et al. Comparison of the oxidation kinetics of different pyrites in the presence of Thiobacillus ferrooxidans or Leptospirillum ferrooxidans[J]. Hydrometallurgy, 1999, 53: 57-72.
[17]Breed A W, Hansford G S. Effect of pH on ferrous-iron oxidation kinetics of Leptospirillum ferrooxidans in continuous culture[J]. Biochemical Engineering, 1999, 13: 193-201.
[18]Dempers C J N, Breed A W, Hansford G S. The kinetics of ferrous-iron oxidation by Acidithiobacillus ferrooxidans and Leptospirillum ferrooxidans: effect of cell maintenance[J]. Biochemical Engineering, 2003, 16: 337-346.
[19]Pesic B, Oilver D J. An electrochemical method of measuring the oxidation rate of ferrous to ferric iron with oxygen in the presence of Thiobacillus ferrooxidans[J]. Biotechnology and Bioengineering, 1989, 33: 428-439.
[20]Ahoned L, Tuovinen O. Microbial oxidation of ferrous iron at low temperature[J]. Appl Environ Microbiol, 1989, 55: 312-316.
������Ŀ: ������Ȼ��ѧ����������Ŀ(50204001); �����ص�����о���չ�ƻ�������Ŀ(2004CB619205-2)
�ո�����: 2005-09-19; ������: 2006-03-10
ͨѶ����: �����, ��ʿ, ������; �绰: 010-62332786; E-mail: lihongxu2001@126.com
[2]�����, ������. ����ұ���е����P������[J]. ��ɫ����, 2003, 55(2): 60-63.
[9]�����, ������, �¾���. ϸ������ļ�����÷���[J]. ��ɫ����, 2003, 55(4): 98- 100.
[10]�����, ������, �¾���, ��. ϸ���������÷���[J]. ��ɫ����, 2003, 55(3): 68-71.
[11]�����, ������. ��ϸ���������̵ĵ绯ѧ(��)[J]. ��ұ, 2003, 12(2): 45-48.
[12]�����, ������. ��ϸ���������̵ĵ绯ѧ(��)[J]. ��ұ, 2003, 12(1): 49-51.
