
���±�ţ�1004-0609(2013)10-3006-09
�����������������Эͬ���û��Ƶ��о���չ
��������ʯ���꣬�� ��������ܣ���ΰ��
(1. ���ϴ�ѧ ��Դ�ӹ������﹤��ѧԺ����ɳ 410083��
2. ���ϴ�ѧ ����ұ��������ص�ʵ���ң���ɳ 410083)
ժ Ҫ��
��������������У���ͬ���͵Ľ��������Ӱ�졢��ٽ�����߸��ԵĴ�л����Ӷ���߽��������ʡ�������������Эͬ���õ�������������������������������������������������������Լ����¾�����¾���Эͬ�����Լ�����֮������û��ƣ�������Эͬ���õ��о�˼·��������������п�����ܽ�;�����ص����Эͬ���õ��о������ؼ���л���P�����á�
�ؼ��ʣ�
����������Эͬ���û�������̬ѧ���������ܻ����������ܽ���
��ͼ����ţ�Q939 �� �� ���ױ�־�룺A
Research development of microorganism synergy mechanisms during bioleaching
YU Run-lan, SHI Li-juan, ZHOU Dan, QIU Guan-zhou, ZENG Wei-min
(1. School of Minerals Processing and Bioengineering, Central South University, Changsha 410083, China;
2. Key Laboratory of Biometallurgy, Ministry of Education, Central South University, Changsha 410083, China)
Abstract: During bioleaching, different types of mixed bacteria interact and cooperate with each other to improve their metabolic activities and increase the metal extracting rate. The synergy types of microorganisms, such as the synergy of iron-oxidizing bacteria and sulfur-oxidizing bacteria, the synergy of autotrophic bacteria and heterotrophic bacteria, the synergy of adsorbed bacteria and free bacteria and the synergy of mesophilic bacteria and thermophilic bacteria and the mechanism of their interactions were reviewed. And the synergy and mineral dissolution ways during bioleaching were discussed, the research methods of the synergy and the roles of important metabolites were discussed.
Key words: leaching microbes; synergy mechanism; ecological methods; functional genes; mineral dissolution
�ҹ������Դ�ḻ����ƶ��ࡢ�����٣���ͳ��ѡұ�ӹ��������ܾ�����Ч�ش�����������Щƶ��ϸ���ӵĿ�����Դ�����⣬��ͳѡұ����������Ը��ӣ���Ҫ��������������ڣ�Ҫ���ϺͶ����豸����Ӧ���ҡ���Ⱦ�Ƚ����ء��ɱ��ϸߡ�����ұ������ָ���ý�������(��Ҫ�ǹž���ϸ�������)����ʯ�е��м۽����ܽ����Һ�У�Ȼ��ͨ����ȡ������ķ�����ȡ�������ӣ��Ʊ��ߴ��Ƚ������ϵĹ��̡��������ܹ���Ч�ش������ֵ�Ʒλ����������ҳɱ��͡������������Ѻ�[1]���ڵ���ȫ���Ʒλ�����Դ�����ȱ������£�����ұ����������Խ��Խ���ԣ�����ȫ��50�������ʵ���˹�ҵӦ��[2]��
�������������ɸѡ�븻���ǿ�չ����ұ������о������̻�Ӧ�õĹؼ���������������࣬�����¶ȵ���Ӧ�̶ȿɻ���Ϊ���¾�(25~42 ��)���ж����Ⱦ�(45~60 ��)�����Ⱦ�(60~80 ��)�ͼ������Ⱦ� (��80 ��)������ͬӪ�����ͻ���Ϊ���������������������������ֿɷ�Ϊ������������������������ͬ����λ��ɷ�Ϊ�����������������Щ��ͬӪ�����͡���ͬ����λ��������ڽ�����ϵ�й�ͬ���ڣ���ͨ������Эͬ������Ч����߽����Ľ������ʺͽ����ʡ�����Acidithiobacillus ferrooxidans (A. ferrooxidans)��Leptospirillum ferrooxidans �� Acidithiobacillus thiooxidans (A. thiooxidans)�����Ի����н���������������ǵ����������¶�Ϊ30~35 �档����A. ferrooxidans��Ҫͨ������Fe2+���������Ҳ���������������Ϳ������ͬʱ�̶�������̼��������[3]�� Leptospirillum ferrooxidansֻ��������Һ�е��������ӣ���������������ԭ���� A. thiooxidans���������������ӣ�ȴ�ܹ������ڵ�����һЩ�����������ϣ������Խ����������в���������������ͬһ������ϵ�У����ǻ�ͨ����ͬ�ķ�ʽ�ĵ��Ӱ�졢�Э��������ܶ��о�����[4]����������A. thiooxidans����ǿ��������A. ferrooxidans�Ľ������á�
�����������������������������ڽ������������ã����õؽ��е�����Ⱥ������ɻ��ܸ�Ч��ɵ�����ת������ǿ���ԵĴ�л����Ӷ���߽����������ʺͽ����ʵ��������û��Ƴ�ΪЭͬ����[5]��ͨ�����ض��������չ����е�����Ⱥ��ṹ��Ⱥ�䶯̬�Լ�����֮�估���������֮�������õ��о������Ը�����˽��������Эͬ���ü���̬���ɣ��Ӷ����ݾ�����о�Ŀ�Ļ�ҵӦ����ѡ����������Ⱥ���Դﵽ�Ż������Ⱥ�ṹ����߽���Ч�ʵ�Ŀ�ġ�
1 ���������Эͬ��������
1.1 ��������������������Эͬ����
��������������У�������������������������Ҫ�Ľ���������Ƿֱ�ͨ������Fe2+ �͵���������������������ͨ������Fe2+����Fe3+��������������ϵ����һ��ǿ���������������������Ӷ�ʹ�������Ӻ����ͷų����������������������������ܳ������������ᣬά������������ϵ�����Ի�����ͨ�����ߵķֹ�Э�����ϴ���ȵ���߽������ʺͽ����ʡ�
GIAVEN��[6]����A. ferrooxidans��A. thiooxidans������ֱ�����˵��������������������Ni3S2���о�����A. thiooxidans��������ʱNi�Ľ���Ũ��Ϊ320 mg/L����A. ferrooxidans��������ʱNi�Ľ���Ũ��Ϊ470 mg/L�������������־���Ͻ���ʱNi�Ľ���Ũ�ȿɴﵽ530 mg/L����ˣ�A. thiooxidans�ļ���������S���������������ƶۻ������ɼ�������Ĥ�ۻ��㣬����֮���Эͬ������������˽��������ʡ�
XIA��[7]Ҳ�о�A. ferrooxidans�� A. thiooxidans֮���Эͬ���ã����Dz��� A. ferrooxidans �� A. thiooxidans������ֱ����п����е�������Ͻ�����ʵ����������A. ferrooxidans����п��Ľ���������A. thiooxidans��ǿ��18 d��п�ĵ��������ʷֱ�Ϊ61%��20%��������Ͻ���ʱ��п�Ľ����ʸߴ�97%�����⣬XRD���������ʾ��A. ferrooxidans����������п��ʱ������������Ҫ��δ��������п�����A. ferrooxidans �� A. thiooxidans��Ͻ���ʱ�������м�ⲻ������ˣ��ƶ�A. thiooxidans���Խ����������������������Ҹ�Ϊ���ף���ȻA. ferrooxidansҲ�������������������Ӻ���ͬʱ����ʱ��A. ferrooxidans��ѡ������������Ϊ��Դ���ʣ����ԣ�A. thiooxidans������������������������á�����A. ferrooxidans �� A. thiooxidans�������ʱ��A. thiooxidans���������������Ķۻ�����ʹ�ý�����Ӧ���Գ�����Ч���У�ͬʱ������������������Ҳ���Լӿ���п��Ľ�������ˣ��������Эͬ�����ڿ����Ч���������з�������Ҫ�����á�
1.2 ����������������Эͬ����
����������Щ������������Щ�Ǽ���������������Щ�������������Ƕ��л�������ó̶��нϴ�IJ�𡣽�����ϵ���л���Ĵ��ڻ�һ���̶�������������������������������������ڣ������л������ۺ�����˥����������϶���л���Ӷ��������������������ԣ����ս��ͽ�������Ч�ʡ�������������Acidiphilium�ļ�������������ͽ�����ϵ���л���ĺ�������������������������ԡ�������Ϊ���������������������Ӱ�����ٽ������Ǿ������������ѱ�������������߽��������ʡ�
GURUNG��[8]�о���������Acidithiobacillus ferrooxidans��������Acidiphilium cryptum �ڽ�����ϵ�е�����á��о�������A. ferrooxidans ����Ч�������ֻ�ѧ��̬���Լ�����Һ�е���������ζ�A. cryptum ���������ƣ���A. cryptum���Խ��������ϵ�еIJ����л���Ӷ���������л����A. ferrooxidans �����������ơ�LIU��[9]�о���������Acidiphilium acidophilum��ת¼ˮƽ�϶�A. ferrooxidans������л��Ӱ�졣�����ʾ���������ʱA. ferrooxidans��ָ�����ӳ�2 d��ϸ�������Ǵ�������5���࣬����Һ���������ӵ�Ũ�������������������йصĻ���������ϵ�(��ͼ1[9])����̶�CO2��صĻ��������Ҳ�ϵ�(��ͼ2[9])���������ʱ��Acidiphilium acidophilum������A. ferrooxidans�е�������������ػ��̶�CO2����ػ���A. ferrooxidans��������л���˼���Ĵٽ����á�
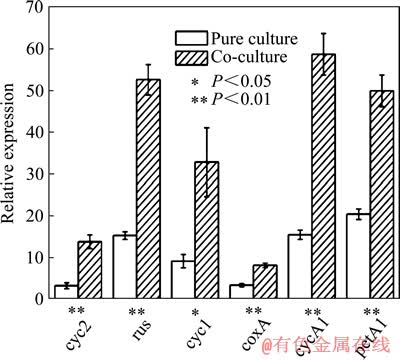
ͼ1 �ڵ�32 hʱA. ferrooxidans�����������������������ػ���������ıȽ�[9]
Fig. 1 Comparison of expression ratio of iron oxidation- related genes of A. ferrooxidans in pure culture and its co-culture with Aph. Acidophilum at 32nd h[9]
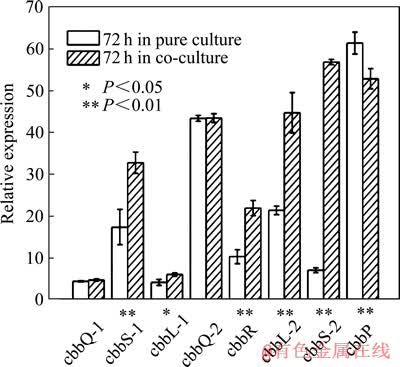
ͼ2 �ڵ�72 hʱA.ferrooxidans���������������CO2�̶���ػ���������ıȽ�[9]
Fig. 2 Comparison of expression ratio of cbb genes of A. ferrooxidans in pure culture and its co-culture with Aph. Acidophilum at 72nd h[9]
1.3 ���������������Эͬ����
����������У�������������λ���Dz�ͬ�ģ��е�ճ���ڿ���ı��棬�е������ڽ���Һ�У����Ƿֱ𱻳�Ϊ�����������������Ȼ�������������ͬʱ�����ڽ�����ϵ�У������Dz����Ĵ�л������ڽ����������ӵ�����ȴ������ͬ��
GAUTIER��[10]������ص��о�������Ϊ�˿�����������������ֱ��ڽ�����ϵ�������ã��ر������һ��װ�ã���250 mL����ƿ���м����һ��Բ���ε�С�ң�С�ҵ��¶���0.1 ��m����Ĥ��ס����ϸ�ṹ��ͼ3[10]��ʵ��a�ֱ���С��A��B��������Sulfolobus metallicus����С��B���˻�ͭ��ʵ��bֻ��С��A������Sulfolobus metallicus����С��B���˻�ͭ��ʵ��c����������ֻͨ�������������������ʵ��b��ͭ�����ʽ���ʵ��a��һ�룬Ҳ����ʵ��c�Ľ����ʸ߳�һ�㣬ԭ������ʵ��b��Sulfolobus metallicus������������ͭ��ı��棬�ڽ�����������ؼ����õ���������Ӳ������ã���Ҫ�����ܽ����Ի�ͭ��Ļ�ѧ���ã�ʹ��ʵ��b��ͭ�Ľ����ʺܵ͡���������һЩ����ʵ�飬��ϸ��������м����ı仯�������ʵ�鲻����Sulfolobus metallicusֻͨ�������������Լ�SO42-��HSO4-��HSO3-��������Sulfolobus metallicus��ͨ�������������Լ�SO42-��HSO4-��HSO3-��SO32-��S2O32-����SO42-��HSO4-�ĺ���Ҫ��û����Sulfolobus metallicus�ĸߺܶࡣ�����ƶ�������Sulfolobus metallicus����������ͭ�����Ķۻ������γ���������Ρ��������κ����������Σ�����������Խ�һ��������Щ���м����õ������κ��������Σ������������ڶۻ����������������Ҫ���ã�ʹ������Ӧ������Ч�ؽ��С�
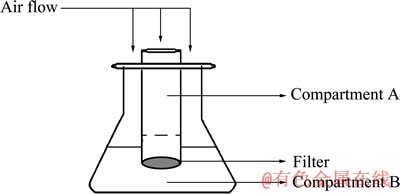
ͼ3 �Ľ����250 mL��ƿװ��ͼ[10]
Fig. 3 Schematic diagram of modified shake flask used in inoculated experiments[10]
�����������ҵӦ���У������������¾����¾���Ͻ��������ڴ��Ͷѽ����к��ڼ��������ȵ��ļ�������һ����нϸߵ��¶ȣ���ͭ��Ľ��������Ӧ���е��¶�һ��Ҳ��60 �����ϣ���ʱ���¾���A. ferrooxidans��L. ferrooxidans��A. thiooxidans��������ֳ�ͽ��������ܵ��ܴ�Ӱ�죬�����¾�ȴ���Է�������Ҫ���á�
��ƽ[11]���������Ⱦ������¾���Ͻ�����Ʒλ��ͭ����о����Խ������ڷ������¾����ܴ������ʵ͵����⡣�������߶��Ի�ͭ��Ϊ���ĵ�Ʒλ��ͭ��ֱ���������Ⱦ������¾������ͻ���������飬��ʯ������80 ��m����Ũ��10%�������ʾ�����¾�����196d����ͭ�Ľ����ʽ�Ϊ22.51%�����¾�������һ��ʱ���������������������ƽ�������Ⱦ����������ڸ��»��������ʯ�еĺ������ʼ����ܳ�����ɽ�����ϵ��pHֵ�����ϳ�ʱ��Ĵ���ȱ仯��������2.0���5.0���ϣ�����ά��ϸ���������������pH��Χ��ϸ���������������½������ڳ������Եġ�ƽ̨�ڡ�������Ч��Ҳ�ϲ�ҽ�������175d��ǰһֱ�����¾��ͣ�����175 d�Ժ����Ⱦ����ֳ�����Ľ������ƣ����ԣ��������Ⱦ���������Ҫ��Լ������ǰ�ڵġ�ƽ̨�ڡ������������������飬���Ⱦ����и��¾����ʹ�ã��ڽ���ǰ���ȼ����¾�ʹ��ʯ�������������в������������֮��������ֽ��ͷ�������ʹ������¶�����ߣ������Ӹ��¾������⣬���¾����������ͷŵ���Դ���ʣ�����������ȣ�����ʹ���ֺ�ĸ��¾��������Ӧ�����������Ӷ���߾��ֻ��ԣ������ڳ�����Ч�Ľ�������ˣ����Ⱦ������¾������ʹ�ã��ȿɱ������¾��Ի�ͭ�����Ч�����ȱ�㣬�ֿɱ������Ⱦ��������ڵġ�ƽ̨�ڡ���ͬʱ�ֿɷ������Ⱦ��Ի�ͭ��Ľ�ǿ����������������ͨ���������־��ĺ������ã����ܳ�ַ��ӳ����Ե����ƣ������Ӱ�졢��ٽ���ʹ�û�ͭ��Ľ������ڽ϶�ʱ�����������Ե���ߡ�
2 ��������ȺЭͬ����
�о����������Эͬ���û��ƣ����ȴӽ����������֣��о������ڸ��ֽ�����ϵ�е�������л���Ⱥ��ṹ����̬�Լ�����ܻ���Ļ��Ա仯�ȣ�������о������������(�������������)�ķֲ��ܽ�;�����ڽ��������в����ĸ����м���������ԭ��������������ܽ�;�����м����ۺ��������ֵ����ݣ�����������������Ⱥ�䶯̬�ı仯����л���Ա仯������Ľ�����Ϊ�������м��������ɼ�ȥ�Ӷ���ʾ��ͬ���͵Ľ��������ڽ�����ϵ�������ӵľ������á���������������ù�ϵ����ͬ���ͽ�������֮���Эͬ���ù�ϵ����һ��������������Эͬ���û��ơ�
2.1 ��������
���������ڸ��������µ�������л������������Эͬ���õ��о�������Ҳ�����о������ϸ����������Ⱥ�ṹ����Ҫ���ܻ���ı�������ȡ���������ѧ�����ķ�չ���о�����������Ⱥ�ṹ��Ⱥ�䶯̬�ķ���Խ��Խ�࣬��ARDRA(Amplified ribosomal DNA restriction analysis)��PCR-RFLP(PCR-Restriction fragment length polymorphism)��Gene-Chip��PCR-DGGE (PCR-Denaturing gradient gel electrophoresis)��Real- time PCR �ȶ���Щ�����ļ�����
���﹦�ܻ�����Ҳ���о���������Эͬ���õĵ���Ҫ�ֶΣ�����ijЩ���ܻ���Ļ��Լ�������Ӱ��������������ܣ��������йؽ������ܻ���ı��������ӳ����Ľ������ܡ�Ŀǰ�����ܻ���������ķ�����Ҫ�й��ܻ���оƬ��Real-time PCR��
���⣬��������������У����������������Ĵ�л����Ҳ��һ���̶���Ӱ���������Эͬ���ã������������(Extracellular polymeric substances, EPS)���ܶ��о��������������ﲻ�����Խ鵼����������������棬�����ܸ�����Һ�е�Fe3+���Ӷ���ǿ����Ч�ʣ���߿���������ʡ�
2.1.1 ����������̬ѧ�о�����
1) �����ݶ�������Ӿ
HE��[12]����DGGE�ķ����о��ڻ�����������������в�ͬ�¶ȺͲ�ͬ��ʼpH������Ⱥ��ṹ��Ӱ�죬ͨ����ȡ��ͬʱ�ڽ���Һ�кͻ�����������������飬���û���16SrDNA��PCR-DGGE�����������Ⱥ���DNA����ָ��ͼ�ף�������������ͳ�Ʒ������н�����DGGE�����������ڳ�ʼpH��ͬ�������£��¶ȶ�����Ⱥ��ṹ��Ӱ��Ƚ����ԡ���30 ���35 �����������£�����Һ��A. ferrooxidans��A. thiooxidans�����ƾ���ͬʱ����35 �������ϵ��Ҳ�ܼ�L. ferrooxidans��������40�����ϵ�У�L. ferrooxidans�Ի�����Ľ������������ã���40��45 ��ʱ��A.caldus��S.thermosulfidooxidans��Ϊ���ƾ������⣬�о�������������������Ⱥ�䶯̬�仯���������Ƶı仯���ɡ�
2) ������DNA����Ƭ������������ø����
ZHOU��[13]�о��ж����Ⱦ��ڽ��跴Ӧ���жԻ�ͭ��Ľ�����������ARDRA�ķ����Խ������̴�����Щ������Щ�����ƾ�������ϸ���졣ͨ��DNA��ȡ��16SrDNA����PCR���������ӡ�ת����һϵ�в�����ܹ��ռ���51�����Կ�¡�ӣ���ø�к������һ����5���������λ(OTUs)���ֱ�ΪZY-1��ZY-2��ZY-3��ZY-4��ZY-5��ͨ��ϵͳ������������ZY-1��ZY-2��ZY-3������L. ferriphilum��ZY-4��ZY-5������A.caldus������ZY-1��ZY-2��ZY-4�ڻ�ͭ��Ľ������������������ã���ռ�����ֱ�Ϊ33.3%��23.5%��29.4%���о������֣�L. ferriphilum��Ⱥܸ�(Լ67%)������Щ��������ڵ�pH���������Ӻ����ߵĻ����У�����о���ͭ��������������ƣ���ʵ��ĺ��ڣ�����Һ�е����Ӹߴ�17.4g/L��pHҲ����1.32���Ӷ��������ڸý�����ϵ��L. ferriphilumռ������λ��ԭ��
3) ʵʱӫ�ⶨ��PCR
ZHANG��[14]����Real-time PCR��������A.caldus��L.ferriphilum��F. thermophilium 3�־��ڻ�ͭ��������������е�Ⱥ�䶯̬�����������ʼ��3�־��Ľ��ֱ���Ϊ1:1:1���������15 dʱ��A.caldus��L.ferriphilum��ϸ�������ϸߣ��ֱ�ԼΪ69%��30%��������28 dʱ��F.thermophilium�������������ӣ�Ϊ22%��A.caldus��L.ferriphilum��ϸ�����������½����ֱ�Ϊ55%��23%��
ARDRA�������Խ�Ϊȷ�ط����������Ⱥ�ṹ�����������Ⱥ��������Ⱥ����Real-time PCR�������ڶ�������ָ����Ⱥ�Ķ�̬�仯����ˣ����ߵĽ�Ͽ�����Ч�ط����������Ⱥ�ṹ��Ⱥ�䶯̬��
2.1.2 �������﹦�ܻ�����Ա仯
��������Ľ�����������ع��ܻ�����ƣ���ͬ��������������Ľ������ܻ�������ͬ�����������ܵIJ�ͬ��������ع��ܻ���ı�������ϣ����Խ������﹦�ܻ�����Ե��о�������Ҫ����Ȼ�������﹦�ܻ����Ѿ����о��ȵ㣬���й�Эͬ���ù��ܻ�����о��ѶȽϴ���صı���Ҳ���١�
������[15]ͨ����ȫ��������ͬ��������������飬ɸѡ�õ����ֲ�ͬ�������Ե�A. ferrooxidans��������ȡ������Ʒ�����ģʽ���������DNA��RNA������ȫ���������оƬ�ӽ����о���������3217�������з�����967������Ϊ��ͬ�ľ��������У������2250��(ռ����������70%)����Ϊ����A. ferrooxidans�����У������֣�320�����������Ծ��������� ���У����м��л(����л)��صĻ���24����������ʡ��ľ��ǡ�����Ǻ�֬���ǵ�����ϳ��뽵����صĻ���33������ⶾ�����йصĻ���15������DNA��л��صĻ���36������������л��صĻ���85������֬����������л��صĻ���5�����뵰���ʵĺϳ���صĻ���26�����������ϳ���صĻ���12������DNAת¼��صĻ���15���������������صĻ���34�������ź�ת����صĻ���10������ת�˺ͽ�ϵ�����صĻ���25�������ԣ�ͨ����������ػ���������ļ�⣬ ���ܶ�����������ܽ��ж����붨����⣬Ҳ�Ϳ���ֱ���˽ⲻͬ����������ʱ�Ľ���Ч����
���ܻ���оƬ�������о��������﹦�ܻ�������һ����Ҫ�ֶΣ����Է�ӳ������Ⱥ�������״���ʹ�л����[16-17]����ʵ�����ڵ�һ��ұ�����﹦�ܻ���оƬ�����ϣ��ɹ������������ڶ���ұ�����﹦�ܻ���оƬ�����Ը�Ϊȷ���ٵط����������﹦�ܻ���������[18]��LIU��[19]���ù��ܻ���оƬ�����о���������(A. thiooxidans)�������������Ⱥ(A. caldus��L. ferriphilum��A. ferrooxidans��S. thermosulfidooxidans��Acidiphilium spp.��F.thermophilum) �������Ե�Ӱ�졣���������A. thiooxidans������ʹ��������(��A. caldus)����������ػ���ı������µ�������������(��L. ferriphilum)����������ػ���ı�����������ߡ����⣬�¾�Ⱥ�����������������ܻ��������Ҳ�ϵ�����ˣ���������(A. thiooxidans)�ļ���������ͭ�������ʺͽ����ʵ���ߡ�
2.1.3 ��������ؼ���л���������������о�
��������(Extracellular polymeric substances��EPS)��ָ������ϸ�������Χ����ϸ����Χ��ˮ������϶��ͨ��䣬�γ�Ģ��״Ĥ�ṹ���������ұ������ճ������Ȼ�л��EPS������Ⱥ���й㷺���ڣ���ϸ����ճ���ۼ�����Ϣ�����������Եȷ���������Ҫ���á�EPS�ijɷֱȽϸ��ӣ���Ҫ�������ǡ������ʡ����ᡢ֬���ᡢ��ȩ�ᣬ�Ұ���ۺ���ɷ�����ֺ�Ӫ����IJ�ͬ����ͬ[20-21]����������������˾��ڴ�л�������������£�����ۺ���ijɷ��Dz�ͬ�ġ���л�������Fe2+��A. ferrooxidans��EPS�к�����ȩ���Fe3+������л���A. ferrooxidans��EPS��ȴû����ȩ���Fe3+��
��ΰ��[22]����AFM�����ͭ����������������������������������Ĺ��̣���ͼ4��ʾ�����������1 d����������������������棬ϸ�������������İ��������γɣ����������3 d���������↑ʼ���ϲ�����������ڰ������ﱾ��֮��ʼ�����������죻���������6 d�����������������������ӣ�������ȫ�������������������Ҹ�����������������ݱ���[23-24]�������������ͨ���������︻����Һ�е�Fe3+���Ӷ����ٿ�����ܽ⡣

ͼ4 ��ͭ���������������������������İ��������AFM��[22]
Fig. 4 AFM images of attached microorganisms on surface of slices of pure chalcopyrite[22]
����������������������е���Ҫ��������Ҫ������[25]��1) �鵼��������������������棻2) ͨ����ȩ������ʸ�����Һ��Fe3+���Ӷ��ڿ�������γ�һ�������ռ������ܽ��������ͼ5[25]��������ԣ���������������EPS��ʾ������ˮ�ı������ʣ���ͨ�����»�����������ˮ����(����)���棻�෴����������������EPS����pH=2�������£������京��0.5%~5%��Fe3+�����γ���ȩ��������ʹϸ����EPS��������磬ͨ���������ý���𤸽��������Ļ�������档Ŀǰ�����ڽ��������������ı������࣬�������������֮������������û�����δ�о���������������о������룬�����������������Ҳ���õ����ơ�
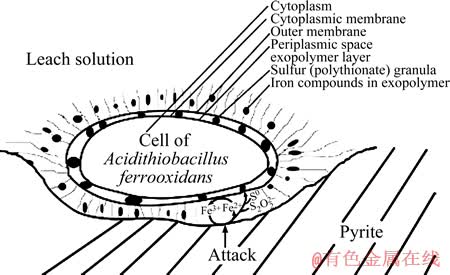
ͼ5 A. ferrooxidansͨ�����������������������û���ģ��[25]
Fig. 5 Contact mechanism model of leaching pyrite by A. ferrooxidans[25]
2.2 ����������������еĿ����ܽ�;��
2.2.1 ������������ܽ�;��
������ʯ�ǽ��������������ú��γɵ��������еĿ�ʯ���ҹ������ϲ��������������еĿ������������Ϊ�����͵�����������������������̶Ȳ�һ����Щ�����ױ�����������Щ�����̶Ƚ��Ƕ������ϸ���绯������أ������������ص㣬��������Ǧп����������մ����˼������ѡ�����һ������������������ߣ����Խ���ijЩ����Ĵ�����ʹ��ʯ�еĽ����ܽ⡣RUSIN[26]������ص��о�����������Ѵ����ĵ������������������Ľ����ʽ�Ϊ1.8%�������ﷴӦ��A��B����Ľ����ʷֱ�Ϊ93.3%��92.4%���Ҷ����ﴦ�����Ŀ�����һ���������Ľ����ʲ������ӣ����������﷽���������Ŀ�ʯ�����ܳ��ס�
����RUSIN[26]�Բ������������������ÿ�����������з��ӵ������в����������Щ�����������Ҳ��̫�˽⣬���ԣ������������������������ϵ�еľ�������û����ϸ������
2.2.2 �����������ܽ�;��
������ͭ��������ұ��Ӧ�õ���Ҫ����֮һ��Ŀǰ��������ͭ��Ĵ��ģ����ѽ��������ȹ��ѵõ��㷺Ӧ�ã��������ҹ���ʯƷλ�����͵IJ��죬Ŀǰֻ���Ͻ�ͭ��ȵ�������ѽ���ҵӦ�õı�����������ͭ����Ҫ������ͭ��ͭ��ͭ���ȣ����ڽ���������ÿһ�ֿ�������÷�ʽ��������ͬ�����м����Ҳ�нϴ���죬���Զ���3����Ҫ�Ĵ�������ܽ�;���ֱ�����ϸ���ܡ�
1) ��ͭ��(Cu2S)
��ͭ�����ԭ����ͭ�������ֽ��پ���ԭ���ö��ɵĴ��������ͭ�ɷָߣ���һ����Ҫ����ͭ��ʯ���ҹ������Ͻ�ɽ�����϶����ȵض��̺��д�����ͭ������[27-28]��������ͭ����������������Ҫ�������������Σ�
Cu2S+Fe2(SO4)3=CuSO4+CuS+2FeSO4 (3)
CuS+Fe2(SO4)3=CuSO4+S+2FeSO4 (4)
����ͭ��ĵ绯ѧ�ܽ��о���������Cu2S��CuS֮�����һϵ���м�������һЩ�����ȶ�����ɺ̶���������ԭ��λ�������ͭ����������������еı仯��绯ѧ�о����һ�£���ʽ(3)�IJ���Ӧ����Cu2-xS������CuS���������ʳ���ҲӦ����һ��������̶ȶ��仯�ġ�Ŀǰ���о�������Դ�����ͺϳɿ�������ڵ�Ʒλ�Ļ�ͭ���ͭ�������Ķ���ѧ�о����Ƚ��١�
2) ��ͭ��(Cu5FeS4)
��ͭ�����ͭ���������У�������Ȼͭ����ȸʯ����ͭ��ͭ���������Ȼ�ͭ�����ͭ������������������ȴ�Ȼ�ͭ��͡��й��������ϰ�ͭ��϶�Ĺ���֮һ����Ҫ�ֲ������϶����ͽ���������ȵء��ܴ���Ϊ6 243��t���������7λ���йر�����ʾ����ͭ����������������ν��У��ֱ����Ӧʽ(5)��(6)��
Cu5FeS4 +8Fe3+��Cu3FeS4 +3Cu2++4Fe2+ (5)
Cu3FeS4 +8Fe3+��3Cu2+ +9Fe2++4S0 (6)
����ͭ��ĵ绯ѧ�ܽ��о�����[29]����ͭ���е��������ܽ⣬����γ�һϵ���м�����Ҫ��Cu8S5��Cu7S4��Cu39S28��CuS��Cu2S���䷴Ӧ���̿��÷�Ӧʽ(7)��(8)��ʾ������������Һ�У�ͭ���ȴӰ�ͭ�����ͷų��������ڽϸߵ�λ�������ڿ���������γ�ͭ����
Cu5FeS4 +3H2O��Cu5S4+Fe(OH)3+3H++3e (7)
Cu5S4+2xH2O��Cu5-xS4+xCu(OH)2+3xH++3xe (8)
3) ͭ��(CuS)
ͭ����һ�ֳɷ�Ϊ��ͭ�Ŀ�����ĺ�ͭ��Ϊ66%����Ҫ����ͭ�Ĵ������︻�����У����ͭ��Ȱ�������ɺ�ͭ�ܸ��Ŀ�ʯ�������ԵIJ��������˹�������IJ������£�ͭ��������������ƿ��÷�Ӧʽ(9)��ʾ��
CuS+Fe2(SO4)3 =CuSO4+S+2FeSO4 (9)
�����������ϵ�У�ͭ���Ǻܶ�������ԭ�����ܽ���м������ױ���[30-31]����ͭ���ڵ绯ѧ�ܽ�����У�����λΪ0.4~0.5 Vʱ����ͭ��缫��������һ������ɫ���ʣ����SEM-EDX���������������ҪΪͭ����
2.2.3 ԭ���������ܽ�;��
��ͭ��(CuFeS2)����Ҫ��ԭ������һ����Ҫ��ͭ����Դ��Լռ�й�70%����ͭ��[32]����ͳ�Ĵӻ�ͭ������ȡͭ�ķ����ǻ�ұ��Ȼ�����չ������ͷŴ�����SO2�����������Ⱦ���������⣬�߶��Ͷ�ʳɱ��Ͳ�������Ҳ��ʹ���ǿ�ʼѰ���������������ͭ������������ʾ���˼�������ơ��ݱ����������Խ����л�ͭ����������������Ҫ��Ϊ����ģʽ��ֱ�����úͼ�����á�ֱ�����á�������÷ֱ���÷�Ӧʽ(10)��(11)��ʾ��
CuFeS2+O2+4H+=Cu2++Fe2++2S0+2H2O (10)
CuFeS2+4Fe3+ =Cu2++5Fe2++2S0 (11)
���У��ڼ��������ϸ��������ֻ������Fe3+�Լ��������������÷�Ӧʽ(12)��(13)����ʾ��
4Fe2++4H++O2=4Fe3++2H2O (12)
S0+3/2O2+H2O=H2SO4 (13)
Ȼ���о�����[33]����ͭ����ܽⲢ�Ǽ�һ����λ�����Ƿֶ���ν��еģ��ܽ�����д��ڵ��м������Ҫ��CuS��Cu2S��CuxS(1��x��2)��CuSn(2��n��8)�ȡ�
��ͭ���ܽ���̵ĵ�һ����������������������ɵ�Cu-S����ľ���ṹ����ֲ��������������������������У�����ͨ��XRD��EDX���ֶ��ڽ��������м�Cu-S����Ĵ��ڡ������������ԭ��1) ��ͭ��������������У�Cu-S�м��������������٣�������XRD��EDX�ķ�����Χ��2) ���ɵ�Cu-S�м�������������������������ڵ������±�������
ACRES��[34-35]����ͬ������XPS�ķ����о���ͭ��ѧ������������̬�仯����������������ѧ����(��pH=1.0�������н���2 h)�����������Ҫ��̬��S0��S22-��SO42-��Sn2-(2��n��8)������Sn2- (2��n��8)��һ�ֶ���������ͬ����ǰ�绯ѧ����������CuxS(1��x��2)��CuxS(1��x��2)����ļ�̬��-2~-1֮��仯����Sn2-(2��n��8)�ļ�̬��-1~0֮��仯�����⣬ͬ������XPS��ͬ������XRD����һ�����ܼ�CuxS(1��x��2)���ʵĴ��ڣ��������ͭ��������������к�������CuxS(1��x��2)���ʡ�
NAVA��[30]��LEE��[36]�Լ�PARKER��[37]�Ի�ͭ��������ܽ�;�����˽�һ���IJ��䡣�����ڵ绯ѧʵ���з��ֻ�ͭ������������������У�1) �ڵ� λ400~600 mV(vs Ag/AgCl)����ͭ������������Cu1-xFe1-yS2 -z(y��x)��2) �ڵ�λ600~700 mV (vs Ag/AgCl)���䣬��ͭ�����������ӿ죬����Cu1-rFe1-sS2-t (r��x, s��y, s��r)���ʡ�������������ɣ���ͭ����������CuS��
������������ͭ����ܽ�;����Ҫ������������ͭ�͵����������Ρ���ͭ����������������ͭҪ���Ķ࣬����ͭ������Cu1-xFe1-yS2-z(y��x)�����ʸ������ǻ�ͭ�������ܽ�ǰ�ڵ����ٲ��衣����ͭ����ɺ�ͭ���������ɵĵ�����Ḳ���ڿ�����棬�γɶۻ�Ĥ���Ӷ�����ͭ�ij����������ڻ�ѧ�����У�������������dz����ѡ�������������У��ں�����������ɼ��������������ᡣ���磬���ж����Ȼ�Ͼ�������ͭ�������һ�������������ᣬ���������˶ۻ����������������ᣬ������ͭ�ij�������ʹͭ�Ľ�����������ߡ�
3 ����
���������ϵͳ�У����ֽ����������ϵ�Ӱ���Эͬ����������֮���Ϊ����������÷�ʽ����Ͼ�Ⱥ�ܸ��õؽ��е�����Ⱥ������ɻ��ܸ�Ч��ɵ�����ת������߸��ԵĴ�л����Ӷ���߽��������ʺͽ������ʡ�ͨ�����û�����ⷽ������Ͻ�Ϊ�Ƚ�����̬ѧ�����Ϳ���ѧ�ֶζ��ض��������չ����е�����Ⱥ��ṹ��Ⱥ�䶯̬�Լ�����֮�䡢���������֮�������õ��о�����һ��������������Эͬ���û��Ƽ�������̬���ɡ�������ѡ����������Ⱥ���Ż������Ⱥ�ṹ��������߿���Ľ���Ч�ʡ�Ȼ�������������һ�����ӵĹ��̣����������͵ķ�Ӧ���绯ѧ��Ӧ���绯ѧ��Ӧ�����ﷴӦ��ԭ��ط�Ӧ�ȡ����ҽ������������û��ܵ��¶ȡ�pH����ʯ���ȡ���Ũ�ȡ�ͨ����������Ũ�ȵȵIJ�ͬ�̶�Ӱ�졣����Ҫ�������˽���õ�������Щ�����������Ҫ����Щ����������������̽����
REFERENCES
[1] RAWLINGS D E. Heavy metal mining using microbes [J]. Annual review of microbiology, 2002, 56: 65-91.
[2] BRIERLEY J A, BRIERLEY C L. Present and future commercial applications of biohydrometallurgy[J]. Hydrometallurgy, 2001, 59(2/3): 233-239.
[3] EECLESTON M, KELLY D P. Oxidation kinetics and chemostat growth of Thiobacillus ferrooxidans on trathionate and thiosulfate[J]. Journal of Bacteriology, 1978, 134(3): 718-727.
[4] LIZAMA H M, SUZUKI I. Bacterial leaching of a sulphide ore by Thiobacillus ferrooxidans and Thiobacillus thiooxidans: ��. Shake flask studies[J]. Biotechnology and Bioengineering, 1988, 32(1): 110-116.
[5] JAIN R, ASHISH P, SREEKRISHNAN T R, DASTIDAR M G. Autoheated thermophilic aerobic sludge digestion and metal bioleaching in a two-stage reactor system[J]. Journal of Environmental Sciences, 2010, 22(2): 230-236.
[6] GIAVENO A, DONATI E. Bioleaching of heazelwoodite by Thiobacillus spp. [J]. Process Biochemistry, 2001, 36(10): 955-962.
[7] XIA Le-xian, LIU Jian-she, XIAO Li, ZENG Jia, LI Ban-mei, GENG Mei-mei, QIU Guan-zhou. Single and cooperative bioleaching of sphalerite by two kinds of bacteria��Acidithiobacillus ferriooxidans and Acidithiobacillus thiooxidans[J]. Transactions of Nonferrous Metals of Society of China, 2008, 18(1): 190-195.
[8] GURUNG A, CHAKRABORTY R. The role of Acidithiobacillus ferrooxidans in alleviating the inhibitory effect of thiosulfate on the growth of acidophilic Acidiphilium species isolated from acid mine drainage samples from Garubathan, India[J]. Canadian Journal of Microbiology, 2009, 55(9): 1040-1048.
[9] LIU Hong-wei, YIN Hua-qun, DAI Yan-xia, DAI Zhi-min, LIU Yi, LI Qian, JIANG Hui-dan, LIU Xue-duan. The co-culture of Acidithiobacillus ferrooxidans and Acidiphilium acidophilum enhances the growth, iron oxidation, and CO2 fixation[J]. Arch Microbiol, 2011, 193(12): 857-866.
[10] GAUTIER V, ESCOBAR B, VARGAS T. Cooperative action of attached and planktonic cells during bioleaching of chalcopyrite with Sulfolobus metallicus at 70 ��[J]. Hydrometallurgy, 2008, 94(1/4): 121-126.
[11] �� ƽ. �и��¾���Ͻ�����ͭ��ϸ����Ⱥ�ṹ����������о�[D]. ����: ����������ѧ, 2008: 3-6.
ZHOU Ping. The research about the bioleaching of chalcopyrite with mesophilic and thermophilic bacteria and the law of bacteria community structure and succession[D]. Kunming: Kunming University of Science and Technology, 2008: 3-6
[12] HE Zhi-guo, YIN Zhen, WANG Xin, ZHONG Hui, SUN Wei. Microbial community changes during the process of pyrite bioleaching[J]. Hydrometallurgy, 2012, 125/126: 81-89.
[13] ZHOU Hong-bo, ZENG Wei-min, YANG Zhi-feng, XIE Ying-jian, QIU Guan-zhou. Bioleaching of chalcopyrite concentrate by a moderately thermophilic culture in a stirred tank reactor[J]. Bioresource Technology, 2009, 100(2): 515-520.
[14] ZHANG R B, WEI M M, JI H G, CHEN X H, QIU G Z, ZHOU H B. Application of real-time PCR to monitor population dynamics of defined mixed cultures of moderate thermophiles involved in bioleaching of chalcopyrite[J]. Applied Microbiology and Biotechnology, 2009, 81(6): 1161-1168.
[15] �� ��, ��ѧ��, �����. ���ڻ���оƬ�������������Ⱥ��ṹ�������������������[J]. ���﹤��ѧ��, 2008, 24(6): 968-974.
SHEN Li, LIU Xue-duan, QIU Guan-zhou. Gene function and microbial community structure in sulfide minerals bioleaching system based on microarray analysis[J]. Chinese Journal of Biotechnology, 2008, 24(6): 968-974.
[16] GILCHRIST D A, FARGO D, ADELMAN K. Using ChIP-chip and ChIP-seq to study the regulation of gene expression: Genome-wide localization studies reveal widespread regulation of transcription elongation[J]. Methods, 2009, 48(4): 398-408.
[17] QIU Guan-zhou, LIU Xue-duan, ZHOU Hong-bo. Microbial community structure and function in sulfide ore bioleaching systems[J]. Transactions of Nonferrous Metals Society of China, 2008, 18(6): 1295-1301.
[18] YIN Hua-qun, CAO Lin-hui, QIU Guan-zhou, ZHOU Ji-zhong, DAI Zhi-min, LIU Xue-duan. Development and evaluation of 50-mer oligonucleotide arrays for detecting microbial populations in acid mine drainages and bioleaching systems[J]. Journal of Microbiological Methods, 2007, 70(1): 165-178.
[19] LIU Yi, YIN Hua-qun, ZENG Wei-min, LIANG Yi-li, LIU Yao, FU Xian, LIU Xue-duan. The effect of the introduction of exogenous strain Acidithiobacillus thiooxidans A01 on functional gene expression, structure and function of indigenous consortium during pyrite bioleaching[J]. Bioresource Technology, 2011, 102(17): 8092-8098.
[20] GUIBAUD G, COMTE S, BORDAS F, DUPUY S, BAUDU M. Comparison of complexation potential of extracellular polymeric substances (EPS), extracted from activated sludges and produced by pure bacteria strains for cadmium, lead and nickel[J]. Chemosphere, 2005, 59(5): 629-638.
[21] HARNEIT K, GOKSEL A, KOCK D, KLOCK J H, GEHRKE T, SAND W. Adhesion to metal sulfide surfaces by cells of Acidithiobacillus ferrooxidans, Acidithiobacillus thiooxidans and Leptospirillum ferrooxidans[J]. Hydrometallurgy, 2006, 83(1/4): 245-254.
[22] ��ΰ��. ��ͭ��������������жۻ�Ĥ���γɻ��Ƽ�����������̽��[D]. ��ɳ: ���ϴ�ѧ��Դ�ӹ������﹤��ѧԺ, 2011: 43-44.
ZENG Wei-min. The formation mechanism of passivation layer and its elimination way during bioleaching of chalcopyrite[D]. Changsha: School of Minerals Processing and Bioengineering, Central South University, 2011: 43-44.
[23] BEECH W B, SUNNER J. Biocorrosion: Towards understanding interactions between biofilms and metals[J]. Current Opinion in Biotechnology, 2004, 15(3): 181-186.
[24] KINZLER K, GEHRKE T, TELEGDI J, SAND W. Bioleaching-a result of interfacial processes caused by extracellular polymeric substances (EPS)[J]. Hydrometallurgy, 2003, 71(1/2): 83-88.
[25] SAND W, GEHRKE T. Extracellular polymeric substances mediate bioleaching/biocorrosion via interfacial processes involving iron (��) ions and acidophilic bacteria[J]. Research in Microbiology, 2006, 157(2): 49-56.
[26] RUSIN P. �����ﻹԭ������Ѵ������������������������Ľ�����[J]. ʪ��ұ��,1995, 53(1): 78-82.
RUSIN P. Improving the extraction percentage of silver and other metals in refractory oxide ore with biological reduction method[J]. Hydrometallurgy, 1995, 53(1): 78-82.
[27] THOMAS G, INGRAHAM T R, MACDONALD R J C. Kinetics of dissolution of synthetic digenite and chalcocite in aqueous acidic ferric sulphate solutions[J]. Canadian Metallurgical Quarterly, 1967, 6(3): 281-292.
[28] ������. ͭ��ʪ��ұ��������ѧ[M]. ����: �й���ѧ����������, 1991: 2.
LIU Chun-peng. Physical chemistry of the copper hydrometallurgy[M]. Beijing: China Science and Technology Press, 1991: 2.
[29] �ƹ���. ��ͭ����ĥ�����֮��ĵ绯ѧ���ü���Կɸ��Ե�Ӱ��[J]. ���������ѡ��, 2009, 46(1): 1-2.
HUANG Guo-zhi. Electrochemical interaction between bornite and grinding media and its effect on the floatability[J]. Metallic Ore Dressing Abroad, 2009, 46(1): 1-2.
[30] NAVA D, GONZALEZ I. Electrochemical characterization ofchemical species formedduring theelectrochemicaltreatment of chalcopyrite in sulfuric acid[J]. Electrochimica Acta, 2006, 51(25): 5295-5303.
[31] MIKHLIN Y L, TOMASHEVICH Y V, ASANOV I P, OKOTRUB A V, VARNEK V A, VYALIKH D V. Spectroscopicandelectrochemical characterizationof the surfacelayers of chalcopyrite (CuFeS2) reacted in acidic solutions[J]. AppliedSurfaceScience, 2004, 225(1/4): 395-409.
[32] DUTRIZAC J E. The kinetics of dissolution of chalcopyrite in ferric ion media[J]. Metallurgical and Materials TransactionsB, 1978, 9(3): 431-439.
[33] SHUEY R T. Semiconducting ore minerals, developments in economic geology series[M]. New York: Elsevier Scientific Publishing company, 1975: 242.
[34] ACRES R G, HARMER S L, BEATTIE D A. Synchrotron XPS studies of solution exposed chalcopyrite, bornite, and heterogeneous chalcopyrite with bornite[J].International Journal of Mineral Processing, 2010, 94(1/2): 43-51.
[35] ACRES R G, HARMER S L, BEATTIE D A. Synchrotron XPS, NEXAFS, and ToF-SIMS studies of solution exposed chalcopyrite and heterogeneous chalcopyrite with pyrite[J]. Minerals Engineering, 2010, 23(11/13): 928-936.
[36] LEE M S, NICOL M J, BASSON P. Cathodic processes in the leaching and electrochemistry of covellite in mixed sulfate-chloride media[J]. Journal of Applied Electrochemistry, 2008, 38(3): 363-369.
[37] PARKER G K, WOODS R, HOPE G A. Raman investigation of chalcopyrite oxidation[J]. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 2008, 318(1/3): 160-168.
(�༭ ����)
������Ŀ��������Ȼ��ѧ�����ѧ������Ŀ(31200382)�������У��������ҵ��������ʦ������Ŀ(1177-721500273)
�ո����ڣ�2012-11-15�������ڣ�2013-06-20
ͨ�����ߣ���ΰ�����ڣ���ʿ���绰��0731-88877472��E-mail: zengweimin1024@gmail.com
ժ Ҫ����������������У���ͬ���͵Ľ��������Ӱ�졢��ٽ�����߸��ԵĴ�л����Ӷ���߽��������ʡ�������������Эͬ���õ�������������������������������������������������������Լ����¾�����¾���Эͬ�����Լ�����֮������û��ƣ�������Эͬ���õ��о�˼·��������������п�����ܽ�;�����ص����Эͬ���õ��о������ؼ���л���P�����á�
[11] �� ƽ. �и��¾���Ͻ�����ͭ��ϸ����Ⱥ�ṹ����������о�[D]. ����: ����������ѧ, 2008: 3-6.
[26] RUSIN P. �����ﻹԭ������Ѵ������������������������Ľ�����[J]. ʪ��ұ��,1995, 53(1): 78-82.
[28] ������. ͭ��ʪ��ұ��������ѧ[M]. ����: �й���ѧ����������, 1991: 2.
[29] �ƹ���. ��ͭ����ĥ�����֮��ĵ绯ѧ���ü���Կɸ��Ե�Ӱ��[J]. ���������ѡ��, 2009, 46(1): 1-2.
