
���������DX1-1������������˾�CMS��
�����ձ����ּ������о�
�����˧, �찮��, ������, ����, �����
(���ϴ�ѧ ��Դ�ӹ������﹤��ѧԺ������ұ��������ص�ʵ���ң����� ��ɳ��410083)
ժ Ҫ��
ժ Ҫ���Դӽ������˻�ͭ������Կ��ˮ�з���õ�������������˾� CMS ����������� DX1-1���������ձ����ּ������о������������ϸ��CMS��DX1-1�����������¶�Ϊ30 �棬����pHֵ�ֱ�Ϊ2.0��3.5��ͨ�������ձ���ͻ������������� DX1-1������������˾� CMS����Ѵ���ʱ��Ϊ60 s����ͻ���ʷֱ�ɴﵽ16.7%��20.0%���ձ������������ DX1-1�ﵽ�ȶ��ڵ�ʱ����ձ�ǰ����20 h�����Ҿ��и���ľ���Ũ�ȣ��ձ�������������˾� CMS����ȫ����������ʱ��Ϊ48 h�����ձ�ǰ��������11 h���ձ���Ͼ������У���ԭ�����չ����ⶨ����30 d��ͭ��������Ũ�ȴﵽ2.78 g/L���������ձ�ǰ�������ͭ��������Ũ��Ϊ2.48 g/L���������30 d����������� DX1-1������������˾� CMS�ľ����������1?1��Ϊ1?20���ҡ�
�ؼ��ʣ�
���������������������˾��������ձ������������
��ͼ����ţ�Q939.97 ���ױ�־�룺A ���±�ţ�1672-7207(2010)02-0393-07
UV-induced mutagenesis and bioleaching of Acidiphilium cryptum and Acidithiobacillus ferrooxidans
YANG Yu, ZHANG Shuai, XU Ai-ling, ZOU Li-hong, LI Li, QIU Guan-zhou
(Key Laboratory of Biometallurgy of Ministry of Education of China, School of Minerals Processing and Bioengineering,
Central South University, Changsha 410083, China)
Abstract: The original strains Acidithiobacillus ferrooxidans CMS and Acidiphilium cryptum DX1-1 isolated from the drainage of some caves rich in chalcopyrite in Dexing in Jiangxi Province of China were studied by UV-induced mutagenesis and bioleaching. The results show that the optimum temperature and pH value are 30 �� and 3.5 for Ac. cryptum DX1-1, and 30 �� and 2.0 for At. ferrooxidans CMS, respectively. After being treated by UV radiating, the optimum UV radiating time of DX1-1 and CMS is 60 s and their positive mutation rates are 16.7% and 20.0%. Ac. cryptum after mutagenesis reaches stationary phase 20 h ahead of the original strain. The most active UV-mutated strain At. ferrooxidans CMS oxidizes all the ferrous ion in 48 h, which is 11 h less than its original strain. After bioleaching with the mixture of UV-mutated strains of At. ferrooxidans CMS and Ac. cryptum DX1-1, 2.78 g/L of copper can be extracted, which can be measured by atomic absorption spectrometry after 30 d, while 2.48 g/L copper can be extracted with the mixture of the original strains before UV-mutation. After bioleaching for 30 d, the proportion of cell density in the cultures of Ac. cryptum DX1-1 and At. ferrooxidans CMS is changed from 1?1 to approximately 1?20.
Key words: Acidiphilium cryptum; Acidithiobacillus ferrooxidans; UV-induced mutagenesis; bioleaching
���������ڼ��˻�����������������������������ѧ����̬ѧ���������ܵ��˹㷺��ע[1]���ر�����������������������ͨ������������Ӧ���ÿ�����������Ӻ����ᣬ�����ǽ���Ӧ�еĹؼ�����[2]����Щ�����ܹ�����������������������������ڻ���������ϸ����ž�[3]������Ϊֹ������ӷ�ˮ�н��õ��˼��������ľ�������������������ϸ��[4-5]������Leptospirillum ferrooxidans, Acidithiobacillus ferrooxidans���ڵ�����[6]���������������Լ����������������У���������������˾�Acidithiobacillus ferrooxidans(At. ferrooxidans)������������Ϊͻ��������Ȼ��������а�����Ҫ��ɫ�����㷺��Ӧ��������ʪ��ұ��[7-8]����Զ��ԣ����Ļ�ԭ��������Ϊ���Լӿ캬��������Ƽ���������������ܽ⡣��������������ϸ�����е������������������������Acidiphilium spp.���þ���������������[9]������Sulfobacillus spp.���þ��Ȳ��������������ֲ��ܻ�ԭ������[10]�����е�Acidiphilium spp.( Ac. cryptum.)����ͨ���컯����ʹ��������ԭ�ɶ�������������������������һЩ���������ɽṹ�����ģ���һЩ�����յ�������[11]��������Ӧ�����������������������Dz����ϵ͡�ԭ���������3�����棺(1) �������ʻ�����(2) ϸ��Ũ�ȵͣ�(3) �����������������б�����[12]����Щ����ؽ����������������еõ������
�ձ�������Ϊһ����Ч����������л����������ֶεõ��˹㷺Ӧ�ã���������һ�ַǵ�����䣬��ʹ���������ʵķ��ӻ�ԭ���е��ڲ��������ܼ�ԾǨ�������ߵ������(��Ϊ����)�����·��ӵ������仯��DNA�����ϵļ����ǿ�����������ߣ�������ऱ���������100���������߷���������DNA���Ķ��ѡ�DNA�����ںͷ��Ӽ�Ľ����������뵰���ʵĽ������Լ�����ऺ�����व�ˮ�����õȣ�������Ҫ�������γ���श����壬�����谭˫���Ľ������ƣ��谭�����������ԣ��Ӷ����»���ͻ�䡣�����Ŵ�ЧӦ��Ҫ������GC��AT ��ת��������ͻ ��[12]����At. ferrooxidans��DNA�У�AT����ĺ���Ϊ46%~47%[13]����Ac. cryptum��AT����ĺ���Ϊ37%~40%�������γ�������श����壬�Ӷ������յ�����ͻ�䡣
1 ���Ϻͷ���
1.1 �������;���
����������ԭʼ����At. ferrooxidans CMS (DQO062118)�� Ac.cryptum DX1-1(DQ529311)�ɽ�������ͭ������Կ�ӷ�ˮ(AMD)�з���õ�[14]������Ȼ�����У�����pH=2.0���¶�Ϊ21 �棬ϸ���ܶ�Ϊ6��109��/L������ͭ���ӵ�����Ũ��Ϊ 0.1 g/L��������2�������������ͭ���������������ͭ��������������
9K�����������ɷ�(����Ũ��)���£�(NH4)2SO4 3 g/L�� KCl 0.1 g/L�� K2HPO4 0.5 g/L��MgSO4��7H2O 0.5 g/L��Ca(NO3)2 0.01 g/L��
CMS����9K������������ʼpHֵΪ2.0 ������4.5% FeSO4��Ϊ��Դ��
DX1-1����9K������������ʼpHֵΪ3.0��������1.5%��������Ϊ��Դ��
9K����������Ϊ9K����������+1%��֬�ۣ�pH=3.5~4.5��
1.2 �������
����ϡ���������������˾���Ũ����ͨ��Ѫϸ����������о��������
1.3 ���������DX1-1������������˾�CMS�������ձ�
�����ڷ����������������������ռ����������ڵľ��壬ϴ�ӣ�����ˮ�Ʊ��ɾ���Һ�������ܶ�NΪ107~108��/mL��ȡ10 mL����Һ����9 cmƽ���У����������������15 W �������30 cm���������䣬����ʱ��ֱ�Ϊ30, 60, 120, 180��240 s������ƽ�ж����顣��ȡ1 mL��Һϡ�ͣ�Ϳ9Kƽ����������ʡ���ȡ1 mL��Һ���ֵ�������0.3% LiCl��9K������������LiCl��������Ϊ����ǿ�ձ�Ч������4 ��ܹⱣ��12 h����30 ��ܹ�����36 h��Ac. cryptum DX1-1����ͻ�����ɴ���ʱ�������At. ferrooxidans CMS����ͻ�������������������ʾ�����At. ferrooxidans CMS����������������ͨ����250 mL��ƿ�����100 mL 9K����������+30 g/L FeSO4��7H2O (6.08 g/L Fe2+)����30 �棬pH=2.0�����������ⶨ���ڳ�ʼ�Σ�ÿ10 hȡ�����Fe2+����Ũ�ȣ����к��ڣ�ÿ3 hȡ�����Fe2+����Ũ�ȡ�
1.4 ����
���������õĿ���ʹ��ǰ�������鲢�ñ�ͪ���Ҵ�ϴ��[15]�������ɷ�(��������)Ϊ��CuFeS2 60.8%��FeS2 20.7 %��CaCO3 8.4%��SiO2 4.6%��
1.5 �����������
��250 mL��ƿ�����100 mL����� 9K����������(������������в���������)������Ϊ5%[16]��ͨ�������ռ�ϸ�������������pH=2.0������ˮϴ��2�Σ�����9K�����������������������������ʣ������¶�Ϊ30 �棬��ʼpH=2.5�����6�������飬ÿ����3��ƽ�����飺(1) �������н����ձ��Ac. cryptum DX1-1��(2) �������н����ձ�ǰAt. ferrooxidans CMS��(3) �������н����ձ��At. ferrooxidans CMS��(4) ���ձ�ǰ2�����������1?1��Ͻ��֣�(5) ���ձ��2�����������1?1��Ͻ��֣�(6) ������(���Ӿ�)��ÿ�־��Ľ�����Ϊ1 mL�������ܶ�Ϊ1��107��/mL���ң� ͬʱ����ҡƿ�ڽ���������
1.6 ��������
ˮ�����ﻯ���ʷ��������ϴ�ѧ�����������IJⶨ������������ԭ�����չ����ⶨ��pHֵ��pH��(pHs-25)�ⶨ��ͭ������30 d�ڵĽ�����ͨ��ԭ�����չ����ⶨ���������ӵ�������ͨ���ظ���ػ�ѧ�ζ����ⶨ��������ͨ��ƽ��������ⶨ��Ac. cryptum DX1-1 ��At. ferrooxidans CMS����ͻ���ʷֱ��ɴ���ʱ����������ӵ������ʾ�����
1.7 ���������ڽ���Һ������Ⱥ����ɷ���
1.7.1 DNA����ȡ�ʹ���
��������Ʒ����Һͨ����Ϊ0.2 ?m�����������������ռ�ϸ����DNA ��ȡʹ��EZ-10 Spin Column Genomic DNA Isolation Kit (Bio Basic Inc) �Լ��У�����ʹ�� Wizard DNA Clean-Up Kit (Promega)�Լ��С�
1.7.2 ����16S rRNA
����2����������16S rRNA����Ƭ�ϣ���������Ϊ27F(5��-AGAGTTTGATCCTGGCTCAG-3��)����������Ϊ1492R(5��-GGTTACCTTGTTACGACTT-3��)��ͨ�����۵���֬��������Ӿ�۲�PCR�����ʹ��Wizard DNA Clean-Up Kit (Promega)�Լ��л���Ŀ��Ƭ�ϡ���16S rRNA����Ƭ�����ӵ�PCR2.1 TOPO�����ϣ�ת����E. coli TOP10F����̬ϸ����(Invitrogen)��ת���������ͿLB��֬ƽ��37 ���ҹ������LB��֬ƽ����백����ù�ء�X-gal��IPTG��ͨ������ɸѡ�������ȡ60����ɫ���䣬ʹ����������M13F(5��-GTAAAACGACGGCCAGTG-3��)��M13R(5��- GGAAACAGCTATGACCATG-3��)����Я������������ϸ���Ʊڣ����о���PCR������
1.7.3 ������Ƭ�ϳ��ȶ�̬�Է���(ARDRA)
16S rRNA PCR�������ᆳHindI��MspI (Fermentas) 37 ��ø�й�ҹ��3.0%��֬��������Ӿ�������������¹۲�ARDRA�����������[17]�н��бȽϷ�����
2 ���������
2.1 ���Կ�ӷ�ˮ����������
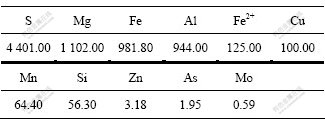
���Կ�ӷ�ˮ�ɷ����1��ʾ���ӱ�1�ɼ����������Ũ��Ϊ4 401.00 mg/L���Ѿ���֤ʵ������ΪAc. cryptum DX1-1����Դ����[15]��Fe3+����ͬ����Ч����Ҳ��Ac. cryptum DX1-1���Դٽ������������Ҫԭ�ɼ�AMD��Ʒʱ��pH=2.0���¶�Ϊ21 �档
��1 ���Կ��ˮ�ɷַ���
Table 1 Composition of AMD ��/(mg?L-1)
2.2 ϸ����������������
pHֵ���¶ȶ�Ac. cryptum DX1-1������Ӱ����ͼ1��ʾ���ɼ�������pH=3.5�����������¶�Ϊ30 �档pH���¶ȶ�At. ferrooxidans CMS������Ӱ����ͼ2��ʾ���ɼ�������pH=2.0�����������¶�Ϊ30 �档����2�־��в�ͬ������pHֵ�Լ�At. ferrooxidans CMS�ڽ���������Ҫ���ã����ԣ��ڽ���������ȡpH=2.5��
2.3 �����ձ���
2.3.1 Ac. cryptum DX1-1�������ձ���
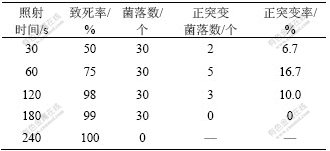
Ac. cryptum DX1-1�������ʺ���ͻ�������2��ʾ���ӱ�2�ɼ���������������ʱ������ȣ���ϸ�����ȶ��ھ��б�ԭʼ�������ľ����ܶ�Ϊ��ͻ��ָ�ꡣ60 sʱ��������Ϊ75%����ͻ����Ϊ16.7%��Ϊ����ձ�ʱ�䡣����ͻ�������ѡȡ����ʱ����̵ľ������ڽ������顣
Ac. cryptum DX1-1�ձ�ǰ���ձ�������������ͼ3��ʾ���ɼ����ձ����ͻ�����ﵽ�ȶ��ڵ�ʱ��Ϊ80 h����ԭʼ������ǰ��20 h�������ܶ�Ҳ��ԭ����109��/mL������1010��/mL��

ͼ1 pHֵ���¶ȶ�Ac. cryptum DX1-1������Ӱ��
Fig.1 Effects of pH and temperature on growth of strain Ac. cryptum DX1-1
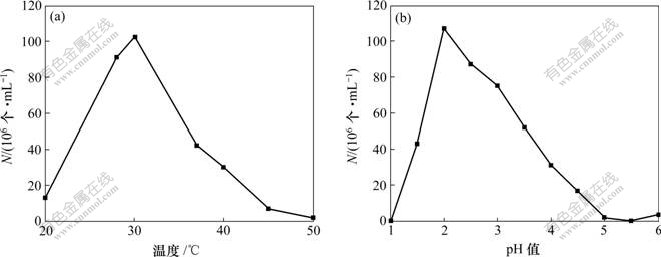
ͼ2 pHֵ���¶ȶ�At. ferrooxidans CMS������Ӱ��
Fig.2 Effects of pH and temperature on growth of strain At. ferrooxidans CMS
��2 �����ձ��Ac. cryptum DX1-1��Ӱ��
Table 2 Effect of UV-induced mutagenesis on Ac. cryptum DX1-1
2.3.2 At. ferrooxidans CMS�������ձ�Ч��
At. ferrooxidans CMS�������ʺ���ͻ�������3��ʾ���ӱ�3�ɼ���At. ferrooxidans CMS��������������ʱ������ȣ���ϸ�����б�ԭʼ���������ߵ�����������Ϊ��ͻ��ָ�꣬60 sʱ��������Ϊ73%����ͻ����Ϊ20.0%����ˣ�60 sΪ����ձ�ʱ�䡣
1���ձ�ǰ��2���ձ��
ͼ3 Ac. cryptum DX1-1�ձ�ǰ���ձ�����������
Fig.3 Growth curves of Ac. cryptum DX1-1 before and after UV-induced mutagenesis
��3 �����ձ��At. ferrooxidans CMS��Ӱ��
Table 3 Effect of UV-induced mutagenesis on
At. ferrooxidans CMS

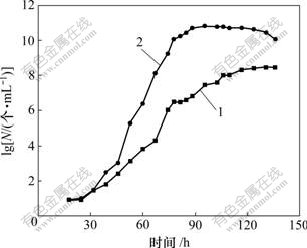
����ͻ�������ѡȡ��������������ǿ�ľ������ڽ������顣At. ferrooxidans CMS�ձ�ǰ�������������������ͼ4��ʾ���ɼ����ձ�����ͻ���������ȫ����������ʱ��Ϊ48 h�����ձ�ǰ���������� 11 h��
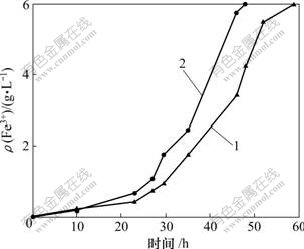
1���ձ�ǰ��2���ձ��
ͼ4 At. ferrooxidans CMS�ձ�ǰ�������������
Fig.4 Fe2+ oxidation rates of strain At. ferrooxidans CMS before and after UV-induced mutagenesis
��ѵ���������ʱ��ȡ����ϸ�����������Լ�����������ԣ��������������At. ferrooxidans CMS �� Ac. cryptum DX1-1�����õͼ����������ձ��Ϊ�ʺϡ���Ϊ�и��������ձ����ɴ��������������ˣ���Щ����Ĺ��ܵò�����ʱ���ͻᵼ�¸�ͻ��� ���������ڵ��±ܹ������£���ø�Ļ����ܵ����ƣ�ϸ���ķ�ֳ�ٶ�ͬ���ܵ������ƣ�ʹ��ͻ���������ʹ�����
2.4 �����������
��ͭ����Ϊ�����Խ��������������������˷�Ӧ�����ܽ�[18-21]��������������ͭ������Һ������ͭ���ӡ����������Ӻ�Ԫ����Ӧ����ʽ���£�
![]()
(1)
At. ferrooxidans�ܹ������ڽ�������в������������ӣ���Ӧʽ����
![]() (2)
(2)
At. ferrooxidans�ܹ������������в���������������Ӧʽ���£�
![]() (3)
(3)
���⣬��������������У�Ҳ�Ե������У���ͭ����������Һ�еĶ���ѧԭ�����£�
![]() (4)
(4)
���ڻ�ѧ����������(ʽ(1))�γɣ����������������(ʽ(3))�б�������
�����Ϸ�Ӧ�в��ѿ�����At. ferrooxidans�����ڷ�Ӧ�ij������С�����������ij�ʼ�Σ���Ҫ��ҡƿ�ڼ�������ʹpHֵά���� 2.5���ң��ṩ��������Ի�����6 d�Ժ�������������Լ�������ˮ���Fe(OH)2+�� ![]() [22]��pHֵ����½���
[22]��pHֵ����½���
��������������У����γɻƼ���������Ļ�����Ϊ�������ڿ�������γ�һ����״�ṹ���谭��ͭ����ܽ�[23]��Acidiphilium���ںܴ�̶��Ͻ���Fe3+-(��)����������Ƽ��������������������������ͻ��Fe2+/Fe3+��������γ�[11]��Acidiphilium�ļ����ܽ��˿���������ܵIJ�״�ṹ��������ͭ���ӵij���������
ѡ��Ac. cryptum DX1-1�� At. ferrooxidans CMS������ͭ�����ͼ5��ʾ���ɼ������������������У�ͭ���ӵĽ�������Ũ�ȳ�����������ǰ15 d��ͭ���ӵĽ�������Ũ�ȿ���������16~30 d������Ũ�������ٶȼ�����ǰ15 d���������ͭ����Ҫ�Σ�ͭ��������Ũ�ȴ����������ͬʱҲ�����˴����ĻƼ���������Ļ����谭�˻�ͭ����ܽ⣬��ˣ�15 d��ͭ���ӵĽ������ڻ������������ձ���Ac. cryptum DX1-1�Ľ�����ϵ�У�30 d��ͭ���ӵ�����Ũ��Ϊ0.52 g/L���������ձ�ǰ��At. ferrooxidans CMS�Ľ�����ϵ�У�30 d��ͭ���ӵ�����Ũ��Ϊ1.82 g/L���ձ�����ϵ��Ϊ1.89 g/L���������ձ�ǰ��At. ferrooxidans CMS��Ac. cryptum DX1-1(������Ϊ1?1)�Ľ�����ϵ�У�����30 d��ͭ���ӵ�����Ũ��Ϊ2.48 g/L���ձ��Ļ����ϵ�У�30 d��ͭ���ӵ�����Ũ�ȴﵽ2.78 g/L�������ձ��������������At. ferrooxidans�������������ԣ����������������
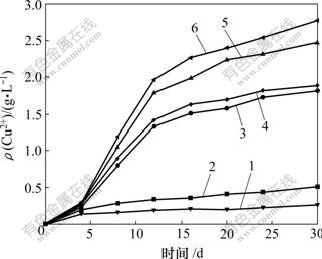
1�������飻2���ձ��Ac. cryptum DX1-1������ϵ��
3���ձ�ǰAt. ferrooxidans CMS������ϵ��
4���ձ��At. ferrooxidans CMS������ϵ��
5���ձ�ǰAt. ferrooxidans CMS��Ac. cryptum DX1-1(1?1)������ϵ��6���ձ��At. ferrooxidans CMS��Ac. cryptum DX1-1(1?1)������ϵ
ͼ5 ��ͬ������ϵCu2+����Ũ��
Fig.5 Cu2+ concentrations extracted in different systems
�����������Կ�����ʹ��2�־���Ͻ����Ч������2�־���������Ч����˵����Ͻ�����һ����ٽ��Ĺ��̡�Ac.cryptum �ӿ��˻�ԭ�ܽ�Ƽ�������������������������At. ferrooxidans�������л�������ٽ���At. ferrooxidans�Ľ�����̡�����Ac. cryptum������������[24]�����ԣ������������Ч����
2.5 ������������Һ������Ⱥ��ṹ����
�����������Ӧ�ij�ʼ�Σ�Ac. cryptum DX1-1��At. ferrooxidans CMS������ͬ��ϸ��Ũ�ȡ�����30 d���ڽ������ձ�ǰ��Ͼ��Ľ�����ϵ�У�����������Ƭ�ϳ��ȶ�̬��(ARDRA)����������Һ��Ac. cryptum DX1-1��At. ferrooxidans CMS��������ԼΪ1?20������ͬ�����£��������ձ���Ͼ��Ľ�������ϵ�У����ǵ�������Ϊ1?18����֤����Ac. cryptum���ǽ�����̵����ƾ���ֻռ����ṹ��С���֣�Э��At. Ferrooxidans��������á���ˣ��������Ժ����������и����������At. Ferrooxidans��������أ����ڽ�����ϵ�н���������Ac. cryptum��
3 ����
(1) ��������ѡ�õ�Ac. cryptum DX1-1 �� At. ferrooxidans CMS�����������¶Ⱦ�Ϊ30 �棬����pHֵ�ֱ�Ϊ3.5��2.0��
(2) �����ձ�Ac. cryptum DX1-1��At. ferrooxidans CMS���ʱ��Ϊ60 s����ͻ���ʷֱ�ﵽ16.7%��20.0%���ձ���Ac. cryptum DX1-1�ﵽ�ȶ��ڵ�ʱ����ձ�ǰ������20 h�����Ҿ��и���ľ���Ũ�ȡ��ձ���At. ferrooxidans CMS���ձ�ǰ���и�ǿ����������������������ȫ����������ʱ��������11 h��
(3) �ձ���At. ferrooxidans CMS ��Ac. cryptum DX1-1(������Ϊ1?1)��Ͻ���30 d��ͭ��������Ũ�ȴﵽ2.78g/L������At. ferrooxidans CMS����������ձ�ǰ��Ͼ��Ľ���Ч����Ac. cryptum���������Ч����
(4) ����30 d��Ac. cryptum DX1-1��At. ferrooxidans CMS����������1?1��Ϊ1?20���ң�Ϊ�Ժ�����������������ṩ�˿�ѧ���ݡ�
�ο����ף�
[1] �ųɹ�, �Ľ���, �����, ��. ��������������˾���������ϵͳ�о���չ[J]. �й���ɫ����ѧ��, 2007, 16(7): 1239-1249.
ZHANG Cheng-gui, XIA Jin-lan, QIU Guan-zhou, et al. Progress in research on Fe2+ oxidation system of Acidithiobacillus ferrooxidans[J]. Transactions of Nonferrous Metals Society of China, 2007, 16(7): 1239-1249.
[2] YIN Hua-qun, QIU Guan-zhou, WANG Dian-zuo, et al. Comparison of microbial communities in three different mine drainages and their efficiency of bioleaching to the low grade of chalcopyrite[J]. Journal of Central South University of Technology, 2007, 14(4): 460-466.
[3] Rawling D E. Heavy metalmining using microbes[J]. Annual Review Microbiology, 2002, 56: 65-91.
[4] Watling H R. The bioleaching of sulphide minerals with emphasis on copper sulphides: A review[J]. Hydrometallurgy, 2006, 84(1/2): 81-108.
[5] Johnson D B, Halberg K B. The microbiology of acidic mine waters[J]. Research in Microbiology, 2003, 154(7): 466-473.
[6] Kelly D P, Wood A P. Reclassi?cation of some species of Thiobacillus to the newly described genera Acidithiobacillus gen..nov., Hallothiobacillus gen.nov. and Thermithiobacillus gen.nov[J]. Evolution Microbiology, 2000, 50: 511-516.
[7] Rawling D E, Silver S. Mining with microbes[J]. Nature Biotechnology, 1995, 13: 773-778.
[8] Rawling D E. The molecular genetics of Thiobacillus ferrooxidans and other mesophilic, acidophilic, chemolithotrophic iron- or sulfur-oxidizing bacteria[J]. Hydrometallurgy, 2001, 59(2/3): 187-201.
[9] Bridge T A M, Johnson D B. Reductive dissolution of ferric ironminerals by Acidiphilium SJH[J]. Geomicrobiology, 2000, 17(3): 193-206.
[10] Bridge T A M, Johnson D B. Reduction of soluble iron and reductive dissolution of ferric iron-containing minerals by moderately thermophilic iron-oxidizing bacteria[J]. Applied Environmental Microbiology, 1998, 64(6): 2181-2190.
[11] Johnson D B, Bridge T A M. Reduction of ferric iron by acidophilic heterotrophic bacteria: Evidence for constitutive and inducible enzyme systems in Acidiphilium spp[J]. Applied Microbiology, 2002, 92(2): 315-321.
[12] ʩ����, ���ɸ�. ��ҵ��������[M]. ����: ������ѧ����������, 1991: 45-48.
SHI Qiao-qin, WU Song-gang. Breeding of industrial microbes[M]. Fuzhou: Fujian Science and Technology Publishing House, 1991: 45-48.
[13] Kulpa C F, Roskey M, Mjoli N. Construction of genomic libraries and induction of iron oxidation in Thiobacillus ferrooxidans[J]. Biotechnology Applied Biochemistry, 1986, 8(4): 330-341.
[14] ����, �찮��. ���Կ��ˮ��һ����Ծ�������ھۺ���ķ��뼰����[J]. �人��ѧѧ��, 2007, 53(6): 753-758.
YANG Yu, XU Ai-ling. Isolation and characteriation of a facultative autotrophic bacterial strain and its cellular polymer granules from acid mine drainage[J]. Transaction of Wuhan University, 2007, 53(6): 753-758.
[15] Mcguire M M, Edwards K J, Banfield J F. Kinetics, surface chemistry, and structural evolution of microbially mediated sulfide mineral dissolution[J]. Geochimica et Cosmochimica Acta, 2001, 65(8): 1243-1258.
[16] ����, ������, ����, ��. һ���ͭ��ר������ϸ���ķ��������[J]. ���ϴ�ѧѧ��: ��Ȼ��ѧ��, 2008, 38(4): 641-644.
YANG Yu, WAN Min-xi, PENG Hong, et al. Isolation and characterization of bacterium for chalcopyrite bioleaching[J]. Journal of Central South University: Science and Technology, 2008, 38(4): 641-644.
[17] YANG Yu, WAN Min-xi, SHI Wu-yang. Bacterial diversity and community structure in acid mine drainage from Dabaoshan mine, China[J]. Aquatic Microbioal Ecology, 2007, 47(2): 141-151.
[18] Rohwerder T, Gehrke T, Kinzler K, et al. Bioleaching review part A: Progress in bioleaching: fundamentals and mechanisms of bacterial metal sulfide oxidation[J]. Applied Microbiology Biotechnology, 2003, 63(3): 239-248.
[19] Suzuki I. Microbial leaching of metals from sulfide minerals[J]. Biotechnology Advances, 2001, 19(2): 119-132.
[20] Schippers A, Sand W. Bacterial leaching of metal sulfides proceeds by two indirect mechanisms via thiosulfate or via polysulfides and sulfur[J]. Applied Environmental Microbiology, 1999, 65(1): 319-321.
[21] Bevilaqua D, Leite A L L C, Garcia O Jr, et al. Oxidation of chalcopyrite by Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans in shake flasks[J]. Process Biochemistry, 2002, 38(4): 587-592.
[22] Sandstorm A, Petersson S. Bioleaching of a complex sulphide ore with moderate thermophilic and extreme thermophilic microorganism[J]. Hydrometallurgy, 1997, 46(1/2): 181-190.
[23] Stott M B, Watling H R, Franzmann P D. The role of iron-hydroxy precipitates in the passivation of chalcopyrite during bioleaching[J]. Minerals Engineering, 2000, 13(10/11): 1117-1127.
[24] Kirsten K. Microbial cycling of iron and sulfur in acidic coal mining lake sediments water[J]. Air and Soil Pollution, 2003, 3(1): 67-90.
