
SBR��������̼ԴŨ�ȶ�����Ӱ��
�����1������Ө1��������1�����ٻ1����漵�2
(1. ������ҵ��ѧ ������ˮ�ʿ�ѧ��ˮ�����ָ������ص�ʵ���ң�������100124��
2. ���������������滮Ժ��������100012)
ժ Ҫ��
A/O)������Ӧ���еľ���Ϊ�о�������SBR���մ���ʵ��������ˮ��̽���ں�����̼��������(m(C)/m(P))�Ծ���(PAOs)�ϳɾۦ��ǻ�����֬(PHB)������Ч����Ӱ�졣�о�������������ں������������л�����Ĵ��ڣ���ʹ����(PAOs)��������PHB�ĺϳɷ�Ӧ�������������գ����������ͷţ����봫ͳ���۲������������λӷ���֬����(UFAs)����Ũ��Ϊ150 mg/Lʱ��PHB�ں����εĺϳ����ʦ�OԶԶ���ڷ�Ӧ��ʼ�������εĺϳ����ʦ�An��ǰ��ԼΪ���ߵ�5����A/O����ģʽ��Ч�ظ�����PAOs������Ч��������ͨ������(OHOs)������������������ԭ��λ(ORP)����ΪA/O����ϵͳ�����������������㡣
�ؼ��ʣ�
��������������������ۦ��ǻ�����֬��������������ˮ��
��ͼ����ţ�X703.1 ���ױ�־�룺A ���±�ţ�1672-7207(2011)10-3220-04
Effect of carbon source on phosphorus uptake in aerobic condition
TANG Xu-guang1, WANG Shu-ying1, PENG Yong-zhen1, ZHANG Jing-qian1, LI Jie-dan2
(1. Key Laboratory of Beijing Water Quality Science and Water Environment Recovery Engineering,
Beijing University of Technology, Beijing 100124, China;
2. Chinese Academy for Environmental Panning, Beijing 100012, China)
Abstract: Aerobic synthesis of poly-��-hydroxybuyrate and uptake of phosphorus in anaerobic/aerobic condition were observed in a sequencing batch reactor (SBR) with real domestic wastewater. The results show that with easily degradable organic substrate, the uptake of ortho-phosphate and the synthesis of PHAs happens insimultaneously, which is consistent with traditional theory. The rate of PHAs synthesized in aerobic condition (��O) is largely higher than the rate of PHAs synthesized in anaerobic condition (��An). And the former is about five times larger than the later. Anaerobic/aerobic condition can enrich the populations of the PAOs, and restrains the growth of the OHOs, availably. ORP profile should be considered as the key characteristic point of phosphorus-uptake in the enhanced bio-logical phosphate removal system.
Key words: biological phosphorus removal; aerobic uptake of phosphorus; PHB; PAO; domestic wastewater
����ǿ������(EBPR)�����ѱ��㷺Ӧ������ˮ�����У��ǽ��ͷ�ˮ�����ŷ����ľ�����Ч�ķ�����EBPR���������ص���Ҫʵ��ϵͳ������/���������Ľ��棬��Ҳ�dz�������(polyphosphate accumulating organisms, PAO)����������ģʽ��������״̬�£�PAO���������û����еĻӷ���֬����(Volatile fatty acids, VFAs)���������������ڵ�����ATP(Adenosine- triphosphate)���ϳ���̼ԴPHB(Poly-��- hydroxybuyrate)���������ATP������ΪADP(Adenosine diphosphate)�����ױ��ͷŵ������У��ں���״̬�£�PHB����ΪATP�������ṩ����������O2���������ջ����е���ʹ����ADP�����������ԴATP���Ա�������������[1-5]�����ǣ�������ʵ��ˮ�Ĵ������л�̼Դ�������٣������ܹ�����PAO������������Ҫ�����ԣ��ܹ���������ε��л����ﺬ���ܵͣ������ε�C/P�ȶԺ�������Ӱ�첢�������ֳ��������ڲ�����ˮ�������У������о�[6-8]��ʾ�����봫ͳ���۲�ͬ�Ľ��������PAO�ں��������п��������л��������PHB�ĸ����������������ͷš����ԣ���EBPR�����У������ε�C/P�ȶ������̵�Ӱ���д���һ���о����ڱ������У�ͨ����Ϊ�ı��ں�������ʱ��C/P�ȣ��ڶ�����-����(A/O)����ѭ���������ĺ������ս����о�����������������������Ρ�������ھۺ����л֮��ı仯�����Լ����Ʋ���ORP�ں����εı仯������з����о���
1 ��������뷽��
1.1 ����װ��
SBR��Ӧ�������л������Ƴɣ��ھ�Ϊ18 cm����Ч�߶�Ϊ65 cm�������ݻ�Ϊ10 L����Ӧ���������װ�ã��ײ���һ��������ͷ�����ÿ���ѹ����������ͨ��ת�������ƿ��������������н�ˮ�����衢��������ˮ�ȹ���ͨ��ʱ��������͵�ŷ�ʵ���Զ��л����������A/O(������-����)��SBR���գ���������Ϊ6 h��ÿ�����ڰ�����˲���ˮ����������(2 h)����������(2~3 h)��������ˮ(1 h)�����ڻ�ˮ��Ϊ8 L����������Ũ��ά����(3 000��200) mg/L��
1.2 ��ˮˮ��
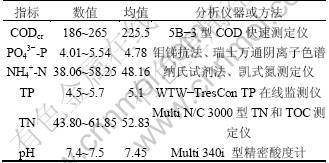
���о�ʹ��ʵ��������ˮ��������β�Ͷ���κ�̼Դ��Ӫ����ȡ�������ˮȡ�Ա�����ҵ��ѧ����С������أ�ˮ��ָ����ʵ�ʳ���������ˮ�Ľ��ƣ�����ʱ�伾����һ���IJ�����ˮ��ָ��(����Ũ��)����1��
��1 ij����С��������ˮ��ˮ��ָ��
Table 1 Characterics of real domestic wastewater mg/L
2 ��������
2.1 �л����ɶ�ϵͳ�ں�����PHAs�ϳɵ�Ӱ��
������ʼVFAs����Ũ��Ϊ0��20.00��149.32 mg/Lʱ�ĸ�����������Ũ�ȱ仯�ֱ���ͼ1~3��ʾ����ͼ1~3���Կ�����������2 h�ڣ�ϵͳ�еı仯��ѭ��ͳ����������ۣ���Ũ����VFAsת��ΪPAOs����PHAs(����PHB��PHV)�Ĺ����������ӡ����У���ǰ1 h�ڣ���������Ϊ6.64 mg/(g��h)��Ϊ��1 h��������2.11 mg/(g��h)��3������������ϵͳ��������1 h��VFA�ĺ�����17.70 mg/L����0.91 mg/L�������������꣬���ں�1 h�е���������Ϊ0.91 mg/L�������õ��л����X���ľ�����������Ҳ����ܵ����ơ�ϵͳ�а��ھۺ���PHB��PHV�ĺ����������������ӣ���2���ں���ĺϳ����ʿ��Կ����������Խ�Ϊ���ȵ����ʽ��кϳɣ��ֱ�Ϊ7.665 mg/(g��h)��3.21 mg/(g��h) �����Կ���ǰ�ߴ�ԼΪ���ߵ�2��������������ʵ��������ˮ�е�VFAs��Ҫ������(HAc)��ɣ�ռ��VFAs��80%���ϣ�����(HPr)������VFAs�ĺ���ֻ���ܺ����ıȵ���20%�����ԣ��ںϳɵľۺ���PHA����Ҫ��PHBΪ������ֻ�ϳ�������PHV����PHB�ĺϳ����������������ܵ��л�����Ũ�ȱ仯��Ӱ�죬�������ں㶨���ʺϳɣ�û����Ϊ����������VFAsŨ�ȵĽ��Ͷ�ʹ�����½���˵��PHB�ĺϳɷ�ӦΪ�㼶��Ӧ���л������Ũ�ȶ���Ӱ���С��
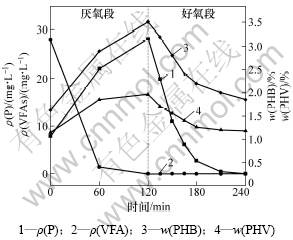
ͼ1 ������ʼVFAs����Ũ��Ϊ0 mg/Lʱ�������ʺ����ı仯
Fig.1 Experimental profiles of different compounds during anaerobic/aerobic cycle when mass concentration of VFAs is 0 mg/L
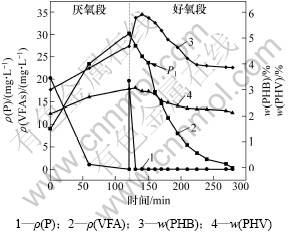
ͼ2 ������ʼVFAs����Ũ��Ϊ20 mg/Lʱ�������ʺ����ı仯
Fig.2 Experimental profiles of different compounds during the anaerobic/aerobic cycle when mass concentration of VFAs is 20 mg/L
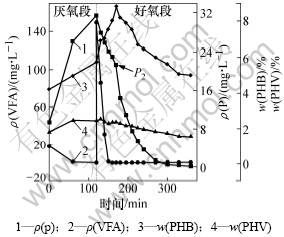
ͼ3 ������ʼVFAs����Ũ��Ϊ149.32 mg/Lʱ�������ʺ����ı仯
Fig.3 Experimental profiles of different compounds during anaerobic/aerobic cycle when mass concentration of VFAs is 149.32 mg/L
�ڽ�������κ��Բ���Ͷ��̼Դ������(NaAc)�ķ������ı������C/P�ȣ�ʹ������ʼVFAs����Ũ�ȷֱ�Ϊ0��20��149.32 mg/L����ͼ1���Կ����������ڽ������״̬���������У������ֽϸߵ���������11.72 mg/(g��h)��������PHB�ĺ���Ҳ��45 min����0.035 mg/(g��h)����0.024 mg/(g��h)��ռ������PHB����������74%��˵��ϵͳ�к�����VFAs����Ũ��Ϊ0 mg/Lʱ����û��PHB�ĸ���������Ҳ�ɷ�����Ч������Ӧ��ʹ��ˮ��Ũ��Ϊ0 mg/L������������ʼVFAs����Ũ�ȵ�����20 mg/Lʱ(��ͼ2)��PHB�ں�������20 min�ڳ��ָ���������0.046 8 mg/(g��h)����0.059 1 mg/(g��h)�������ڴ˽��ദ������״̬���������������Ե���ǰ�ߣ����ҿɿ����������յ�P1��ǰ�������������ܴ����ԣ���������ʼVFAs����Ũ��������149.32 mg/Lʱ����������������ԡ���ͼ3���Կ���������VFAs�������轵���ں�������40 minʱ�京������Ϊ0����������Ϊ89.59 mg/(g��h)������VFAs��ת��;������2��������Ӧ��
(1) OHO�ں���״̬�¶�VFAs�Ľ��⣬��Ҫ����OHO����������������ã����ǣ����������������ڸ�ϵͳ�У����ڳ��ڵ�����/������������ģʽ��OHO�Ļ�Ա����ƣ��ں����ζ�VFAs�Ľ���� ���ޡ�
(2) PAO��VFAs��ת�����档�ڶ�ϵͳ��PHB�ļ�ⷢ�֣����ں������ڼ�VFAs�轵�Ĺ��������д������ɣ����Һϳ����ʺܸߣ�Ϊ36.97 mg/(g��h)���ɴ˿��Կ������ں��������£�PHB�Կ��Ա��ֽϸ����ʽ��кϳɣ����б��ڴ�ͳEBPR�����к���PHB�������ۡ�����Ũ�ȱ仯���Կ�������ͼ3��������������ʹյ�P2���ڸùյ�ǰ������������ͼ2�е�һ�����ܴ��ϵͳ��PHB�仯���߿��Կ��������յ�P1��P2��Ӧ���Ǻ���PHB�ϳɵĽ������������ں���״̬�¿��Ը��ٵ����û����е��л��������ͬ�����ã������ںܴ�̶���Ӱ���˺�������Ӧ��
���⣬���Դ������κͺ����ε�λ�����кϳɵ�PHB��(��)���Ƚ���2��״̬��PAO�Ķ��л����������������ڴ˽������¼��裺
(1) �������Σ�VFAsȫ������PAOs����PHB�ĺϳɡ�
(2) �ں����Σ�VFAs��ת��ȫ������2��������Ӧ�����У�һ��ΪOHO�ĺ������⡣���ǣ��������������������е�ϵͳ�У�PAO�������ξͿ�ʼ�����л������������ϳ����ã�������������������PHA�����ڴ˹����в���ATP��ά�����������IJ������������沢���ڽ�ǿ����״̬����Ҳʹ��PAO������/������������ϵͳ�о�������������д������������������о�����������ʹ�������������ں����εò�����Ч��������ʹ������������Ϊϵͳ�����ƾ�Ⱥ�����ԣ��ڳ��ڵ����������������е�ϵͳ�У�PAO����������ռ�����ƣ���ͨ����������PAO��Ӱ���С��������Ӱ���ں����ζ������л�����ľ�����
��һ��ΪPAO����PHB�ĺϳ�Ϊ��Ҫת��VFAs�ķ�Ӧ���������Σ���AnΪ7.665 mg/(g��h)�����ں����Σ���OΪ36.97 mg/(g��h)����O��ԼΪ��An��5����PAO�ں��������µĺϳ��������Ա�������������PHB��ǿ������˵���ں��������е����㹻��VFAs����ʱ��PAOs�������������PHAs�ĺϳɣ������������Ը�������״̬�µĺϳ����ʡ������ԭ���д���һ���о���
2.2 A/O����ģʽ��ϵͳ�о�Ⱥ��Ӱ�����
�ӹ�����������ˮ���ﴦ��ϵͳ�е�������Ҫ�а�������(AOB)����������������(NOB)������(PAO)��һ�������������(OHO)�ȣ����ڳ���ϵͳ������Ҫ���õľ�Ⱥֻ��PAO��������Ⱥ��������������Ӱ�쵽ϵͳ�ij���Ч�ܡ��ڸ�SBRϵͳ�У���ȡ����/������������ģʽ��ϵͳ�еľ�Ⱥ�����������У���ϵͳ�����������У�������������AOB��NOB����ϴ��ϵͳ����ʹ�ں����μ���û���������õķ��������Ƕ�������͵���(ϵͳ�����¶�ά����13 ������)���¡��ڵ������������������������������������ҽϳ�ʱ��ʹ����ϵͳ�еò�������������ͨ��ÿ���ڵ�����ܿ챻��ϴ��ȥ����Σ�����ϵͳ��Ͷ��������ˮ�����Ƚ��������Σ�����ʱPAO�ܹ������������г������ԭˮ�е��л�����������������ڣ���Ҫ��ѭ�����ա����桪���á�������ģʽ����������Ũ�Ƚϸ�ʱ�����ォ���ʵIJ��ֻ�ȫ�����ȿ������գ������ھۺ������ʽ���棬Ȼ������ȱ��ʱ�����ô���ľۺ��������ֳ[9-10]�����ǣ�����һ�����������˵����������ģʽ�����������ã������ԡ�ֱ�����á���ֳģʽΪ��������Щ��Ⱥֻ���ں��������в����д����������ʱ�ſ��������������ɼ������PAO������������˵��OHO���������ܵ����ƣ��������ʻ��������ԣ��ڸ�ϵͳ�У�ͨ��A/O����ģʽ����������Ч�ظ����˶Գ����й��ľ�ȺPAO����������������������������OHO�ķ�Ӧ���ԡ�
2.3 ��ͬC/P����VORP�ĺ���ָʾ����
�����������о������������������Ŀ���һ����ORPָ��VORP����ӳ����VORPΪ��ֵʱ�����������ף���VORPΪ��ֵʱ������ֵԽ�ߣ�������������Խǿ��һ����Ϊ��Ӧ��VORP������-200~-300 mV���Ժ�����Ϊ������ȵط��Ӿ������������ã������ṩ�㹻������DO������������������PHB���н���ʱ��������Ϊ���յ����������������ʵ������ȵ�ת��PHB���ͷų��㹻��ATP����������������衣һ��Ӧ���˶ε�DO����Ũ�ȿ�����2 mg/L���ϡ����ǣ���VORP���ں��������еĿ��Ʋ���������ڸ��о��У��ڲ�ͬ������C/P�������·����˲�ͬ��VORP�仯���ɣ����ܹ�ָʾ����Ӧ ���̡�
ѡ������γ�ʼVFAs����Ũ�ȷֱ�Ϊ0��20.00��149.32 mg/Lʱ�ĵ��������ڵ�VORP�仯���ɣ���ͼ4���������������ڷ�Ӧ�տ�ʼ��5~10 min�ڣ�VORPѸ���ɿ�ʼ��-300 mV������-375 mV���ң��������ڴ�����������ˮ��������DOѸ�ٺľ����Լ���������̬��(NOx-)���ϱ���ԭ�ɵ���������������Ӧ���е�������ԭ��λ���Ͻ��ͣ����������������������Ľ��У���̬�����ϼ��٣���������Ӧ���У�������ԭ״̬�ı仯���練��������������ԭ״̬�ı仯���ȴ����ԣ�VORP�ı仯Խ��ԽС��������������ʱVORPѸ���½���������������Ϊһ�յ�(��ͼ��D)����һ��ָʾ��ϵͳȱ���������̵Ľ���������̬����ʧ��ϵͳ��������״̬�����������ж�ϵͳ�����������Ρ�
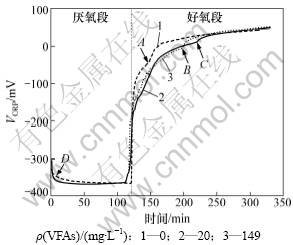
ͼ4 ��ͬ�л�����ORP�仯���
Fig.4 Experimental profiles of ORP during the anaerobic/aerobic cycle at different OLR
�ڽ����������貢�ֱ�����ϵͳ�л����ɺ�ϵͳ����������Σ��ڴ˹�����VORP�ı仯Ϊ���ں����εĿ�ʼ10 min�ڣ���������DO����Ũ��Ѹ�����ߣ���ʱVORP���ߵ�����Ҳ�ܴ���-375 mVѸ��������-100 mV���ڴ�֮��������Σ�3�ֲ�ͬ�л������ں����ε�VORP�仯���ֲ��죬�ھ�������VORPѸ������֮������VFAs����Ũ��Ϊ0 mg/L��ϵͳ���ȴﵽ�յ�A����ʱ��̣�˵����C/P��Ϊ0ʱ��������ں���������������������PHAs���������������Ӧ��ʹ�������ʱ����ڽϸ�ˮƽ,���ں�����ʼVFAs����Ũ�ȷֱ�Ϊ20 mg/L��149 mg/L��ϵͳ�ڣ�VORP�������������ﵽ�յ�B��C����������֪��������ʼ�ĸ�C/P�Ȼ�Ծ�����������������Ӱ�죬����ڵ���ḻ�������¼����ں��������������л��������PHAs�ĺϳɣ���Ӱ�쵽�������ʣ��Ӷ��������յ���ͺ�
����������VORP��Ϊ���Ʋ������о��У��ཫ����Ϊ���������������������Ա���ʵ�ֶ̳��� �� [11-13]���������ڷ�������������������������ȱ����������ָʾ����[14-15]����û�еó�VORP�ں����������еı仯���̼�������ָʾ���á������ڸ������У���A/O�������е�SBRϵͳ�У��ں�������������Ҫ�������õ��Ǿ�������PAOs��OHO�Ļ��Ա��������ƣ����ԣ�VORP�ı仯������Чָʾ��������̼Դ��PHB�ϳɽ�����ϵͳ��ʼ����������Ρ�
3 ����
(1) ��3�ֲ�ͬ������C/P�������£�ϵͳ���ֳ��б��ڴ�ͳ������������PHAs�ĺ�������������ͬʱ���У�����������PHB�ϳ����ʦ�AnΪ7.665 mg/(g��h)�����ں����Φ�OΪ36.97 mg/(g��h)����O��ԼΪ��An��5����PAO�ں��������µĺϳ��������Ա�������������PHB�ĺϳ�����ǿ������˵���ں��������У������㹻��VFAs����ʱ��PAO�������������PHA�ĺϳɣ������������Ը�����������״̬�µĺϳ����ʡ�
(2) ������ʵ��������ˮ��SBRϵͳ�У�ͨ��A/O����ģʽ����������Ч�ظ����˶Գ����й��ľ�ȺPAO����������������������������OHO�ķ�Ӧ���ԡ�
(3) �ڸ������У���A/O�������е�SBRϵͳ�У���������Ҫ������Ӧ������Ⱥ��ΪPAO������ ORP�Ĺյ�����Чָʾ��PHB�ϳɵĽ����κ�ϵͳ��ʼ����Ŀ������Ρ�
�ο����ף�
[1] Satoh H, Mino T, Matsuo T. Uptake of organic substrates and accumulation of poly-hydroxyalkanoates linked with glycolysis of intracellular carbohydrates under anaerobic conditions in the biological excess phosphate removal processes[J]. Water Sci Technol, 1992, 26(5/6): 933-942.
[2] Smolders G J F, van der Meij J, van Loosdrecht M C M, et al. Model of the anaerobic metabolism of the biological phosphorus removal process: Stoichiometry and pH influence[J]. Biotechnol Bioeng, 1994, 43(6): 461-470.
[3] Smolders G J F, van der Meij J, van Loosdrecht M C M, et al Stoichiometric model of the aerobic metabolism of the biological phosphorus removal process[J]. Biotechnol Bioeng, 1994, 44(7): 837-848.
[4] Pereira H, Lemos P C, Reis M A M, et al. Model for carbon metabolism in biological phosphorus removal processes based on in vivo 13 C-NMR[J]. Water Res, 1996, 30(9): 2128-2138.
[5] Smolders G J F, van der Meij J, van Loosdrecht M C M, et al. A structured metabolic model for anaerobic and aerobic stoichiometry and kinetics of the biological phosphorous removal process[J]. Biotechnol Bioeng, 1995, 47(3): 277-287.
[6] Brdjanovic D, Slamet A, van Loosdrecht M C M, et al. Impact of excessive aeration on biological phosphorous removal from wastewater[J]. Water Res, 1998, 32(1): 200-208.
[7] Guisasola A, Pijuan M, Baeza J A, et al. Aerobic phosphorus release linked to acetate uptake in bio-P sludge: Process modeling using oxygen uptake rate[J]. Biotechnol Bioeng, 2004, 85(7): 722-733.
[8] Ahn J, Daidou T, Tsuneda S, et al. Transformation of phosphorus and relevant intracellular compounds by a phosphorus accumulating enrichment culture in the presence of both the electron acceptor and electron donor[J]. Biotechnol Bioeng, 2002, 79(1): 83-93.
[9] van Loosdrecht M C M, Pot M A, Heijnen J J. Importance of bacterial storage polymers in bioprocesses[J]. Water Science and Technology, 1997, 35(1): 41-47.
[10] Majone M, Assanisso P M, Ramadori R. Comparison of carbon storage under aerobic and anoxic conditions[J]. Water Science and Technology, 1998, 38(8/9): 77-84.
[11] �߾���, ������, ����Ө. DO��ORP��SBR����������������ع�ϵ[J]. ������������ѧѧ��, 2002, 35(1): 61-65.
GAO Jing-feng, PENG Yong-zhen, WANG Shu-ying. Relationships between DO, ORP and nitrification, denitrification in SBR processes[J]. Journal of Harbin University of Civil Engineering and Architecture, 2002, 35(1): 61-65.
[12] �ߴ���, ������, ֣����. SBR�����ж̳������������Ĺ��̿���[J]. �й���ˮ��ˮ, 2002, 18(11): 13-18.
GAO Da-wen, PENG Yong-zhen, ZHENG Qing-zhu. Process control of shortcut nitrification-denitrification in sequencing batch reactor process[J]. China Water & Wastewater, 2002, 18(11): 13-18.
[13] Chang C H, Hao O J. Sequencing batch reactor system for nutrient removal: ORP and pH profiles[J]. Journal of Chemical Technology and Biotechnology, 1996, 67(1): 27-38.
[14] ������, ������, ����Ө, ��. ̼Դ����̬��Ũ�ȶԷ���������Ӱ�켰ORP�ı仯����[J]. ������ѧ, 2004, 25(4): 54-58.
WANG Ya-yi, PENG Yong-zhen, WANG Shu-ying, et al. Effect of carbon source and nitrate concentration on denitrifying dephosphorus removal and variation of ORP[J]. Environmental Science, 2004, 25(4): 54-58.
[15] Schon G, Geywitz S, Mertens F. Influence of dissolved oxygen and oxidation-reduction potential on phosphorus release and uptake by activated sludge from sewage plants with enhanced biological phosphorus removal[J]. Wat Res, 1993, 27(3): 349-354.
(�༭ �²ӻ�)
�ո����ڣ�2010-11-15�������ڣ�2011-01-28
������Ŀ�����ҡ�ʮһ�塱 �ش�Ƽ�ר��(2008ZX07209-003)�������н�ί�Ƽ�����ƽ̨��Ŀ(PXM2008_014204_050843)
ͨ�����ߣ������(1982-)���У��ӱ���̨�ˣ���ʿ�о���������ˮ��Ⱦ���ƹ��̼�ˮ�����ָ��о����绰��010-67392627��E-mail��txg@emails.bjut.edu.cn
ժҪ��������/����(A/O)������Ӧ���еľ���Ϊ�о�������SBR���մ���ʵ��������ˮ��̽���ں�����̼��������(m(C)/m(P))�Ծ���(PAOs)�ϳɾۦ��ǻ�����֬(PHB)������Ч����Ӱ�졣�о�������������ں������������л�����Ĵ��ڣ���ʹ����(PAOs)��������PHB�ĺϳɷ�Ӧ�������������գ����������ͷţ����봫ͳ���۲������������λӷ���֬����(UFAs)����Ũ��Ϊ150 mg/Lʱ��PHB�ں����εĺϳ����ʦ�OԶԶ���ڷ�Ӧ��ʼ�������εĺϳ����ʦ�An��ǰ��ԼΪ���ߵ�5����A/O����ģʽ��Ч�ظ�����PAOs������Ч��������ͨ������(OHOs)������������������ԭ��λ(ORP)����ΪA/O����ϵͳ�����������������㡣
