
3��ϸ������ľ���ص����������о�
�Ż�1������Ԫ1����ӽ��2����Ծ��1����Ƚ2
(1. ���ϴ�ѧ ұ���ѧ�빤��ѧԺ������ ��ɳ��410083��
2. ���ϴ�ѧ ������ѧ�뼼���о�Ժ������ ��ɳ��410082)
ժ Ҫ��
B����ƽ��Դ��������ʴ���з���õ���3��ϸ��Acinetobacter sp. B-2��Pandoraea sp. B-6��Novosphingobium sp. B-7������ɫ���飬������3��ϸ����Һ�����������µ�ľ���ؽ������ܣ�������ľ���ؽ��������ĵ��ؽ����о�������ȷ����3������˵Ľ����������о������������3��ϸ������ʹ��������RB����Ⱦ����ɫ���������ܹ���ľ���ؽ���ø����3��ϸ����ľ���ؽ������ʽϿ죬��5��ľ���صĽ�����������ȶ����������˵Ľ����������£���Acinetobacter sp. B-2����ԴΪ����泥���ԴŨ��Ϊ0.01 mol/L�������¶�Ϊ30 �棬��ʼpH=7.0��ҡ��ת��Ϊ120 r/min����Pandoraea sp. B-6����ԴΪ�������泥���ԴŨ��Ϊ0.03 mol/L�������¶�Ϊ30 �棬��ʼpH=7.0��ҡ��ת��Ϊ120 r/min����Novosphingobium sp. B-7����ԴΪ����泥���ԴŨ��Ϊ0.01 mol/L�������¶�Ϊ30 �棬��ʼpHΪ5.0��ҡ��ת��Ϊ120 r/min�������˵Ľ��������£���3��ϸ����3���ľ���ؽ����ʾ��ɴﵽ30%~35%��
�ؼ��ʣ�
ľ������ľ���ؽ���ø���������������������
��ͼ����ţ�X172 ���ױ�־�룺A ���±�ţ�1672-7207(2011)10-2922-07
Cultural condition regulations for three lignin degrading bacteria
ZHANG Huan1, CHAI Li-yuan1, ZHU Yong-hua2, CHEN Yue-hui1, JIN Ran2
(1. School of Metallurgical Science and Engineering, Central South University, Changsha 410083, China;
2. Institute of Life Science and Biotechnology, Hunan University, Changsha 410082, China)
Abstract: The decolorizing reactions of three bacteria were studied through Azure-B and Remazol Brilliant Blue dye plate, which were isolated from erosive bamboo slips of Kingdom Wu, identified as Acinetobacter sp. B-2, Pandoraea sp. B-6 and Novosphingobium sp. B-7, respectively. The lignin degrading characteristics of three bacteria were investigated in the liquid medium. The effects of culture conditions on lignin degrading ability of the three bacteria were also studied, and the appropriate culture conditions were determined. The results show that three bacteria can decolorize Azure-B and Remazol Brilliant Blue dye and produce ligninolytic enzyme. Three bacteria can degrade lignin very well with a relatively high degrading rate, and the plateau of degradation is observed after 5 d incubation. Appropriate culture conditions for Acinetobacter sp. B-2 are achieved as follows: NH4NO3 as nitrogen source, 0.01 mol/L of nitrogen source concentration, 30 �� of temperature, the initial pH=7.0, 120 r/min of rotation speed. The appropriate culture conditions for Pandoraea sp. B-6 and Novosphingobium sp. B-7 are similar to that for Acinetobacter sp. B-2, and NH4H2PO4 as nitrogen source and 0.03 mol/L of nitrogen source concentration for Pandoraea sp. B-6, the initial pH=5.0 for Novosphingobium sp. B-7, respectively. In addition, the degrading rates of three bacteria can reach 30%-35% after 3 d incubation under appropriate degrading conditions.
Key word: lignin; ligninolytic enzyme; degradation conditions; bamboo slips of Kingdom Wu
ľ��������Ȼ���к�����Ϊ�ḻ��һ����Ȼ�л��߷��Ӿۺ�����㷺������ֲ��ϸ�����У�����ά�غͰ���ά��һ��ֲ���֧�ŹǼܡ�������ϳɵ�Ӱ�죬ľ���ط��Ӳ�����ά���������е�һ�����ʽ�������Ա�����������Ϊ�ṹ��Ԫ��ͨ��̼̼�����������������Ѽ������Ӷ��ɵ�������ά�ռ���״������߷��Ӿۺ�����������������ӣ�ľ���ز���������Ȼ�߾���(��ά�ء����۵�)һ���ױ����⣬��Ŀǰ���ϵ������ѽ���ķ����廯����֮һ[1]��ľ���ص����オ�ⲻ��ȡ�������������Ľ��������������ںܴ�̶�����������Χ�Ļ���������Ŀǰ�������й�����ľ���ؽ���Ӫ�����ػ��Ƶ��о�������ľ���ؽ���ø������̼Դ����Դ��̼���ȡ�pH���ܽ���Ũ�Ⱥͽ������ӵ�����Ӱ�졣Kirk��[2]ͨ���о�ľ���ؽ���ģʽ������ԭëƽ���(Phanerochaete chrysosporium)��Ϊ����ԴŨ�ȶ���ľ���ؽ����Ӱ���Ϊ�������͵�������ľ���صĽ��⣻���������������˿����γ���������ľ���صĽ��⣻ľ���ؽ�������pHΪ4.0~4.5��������[3]����ɸѡ����1������F2��ľ���ؽ���ø�������Խ������о��������������̼����������������LiPø�IJ�������̼ԴŨ�ȶ����MnP��Ӱ�첻�������������¶�������MnP�IJ������������ľ���ؽ���������Ҫ���������ľ���ؽ��⡢��ø�����Ż���ʵ��Ӧ�õȷ�����о���Ŀǰ���\��Ӧ�÷�����ȵ����[4-5]��Ȼ����ϸ����������Ѹ�١���Դ�㷺�����ڻ��̸��켰��ҵ�����������ƣ���ˣ���ϸ��ľ���ؽ�����о�Ҳ���кܴ��Ӧ�ü�ֵ��Ϊ�ˣ���������ǰ�ڴ��������ʴ�������з��롢ɸѡ�õ�3�����ľ���ؽ���������ϸ��Acinetobacter sp. B-2��Pandoraea sp. B-6��Novosphingobium sp. B-7[6]��Ŀǰ�ѷ��ֲ����˾���Acinetobacter��һЩ������н���ľ���ؼ�����ػ������������Delneri��[7]�о���Acinetobacter calcoaceticus DSM 586�����ľ���ص��廯����Ľ����л�����ָþ�������Ч�ؽ��ⷴʽ��κ��Ͷ��DZ���ϩ��,���Խ��ⷴӦ���ƽ����˷�����Vasudevan��[8]��ɭ�������з���ɸѡ�õ���1��Acinetobacter sp.���ⶨ���꽵��14C��ǵ�����߷��Ӿۺ���(DPH�����㷺������ľ���ص�ģ�ͻ�����)����ľľ����CO2�ۻ���������6 d�ֱ�ɴ�38.9%��27.2%����Ȼ��Pandoraea����Novosphingobium����ϸ������ľ���ؼ���ľ������ػ�����ı������٣�����֤�ݱ��������ܽ��ⷼ���ࡢ�������л��Pandoraea sp. �ܽ����л���ɱ�����[9]����Novosphingobium sp. �ܽ��������������ȷӵȶ�����������[10]��Ŀǰ�����Ƕ���3��ϸ����ľ���ؽ����������ص��о����١�Ϊ�ˣ��������߲���������ˮ��ľ���ػ�������Ϊ��Ȼľ���ص�����̽�ֵ�Դ����ԴŨ�ȡ��¶ȡ�pH��ҡ��ת�ٵ����ضԾ���ľ���ؽ������ܵ�Ӱ�죬�Ӷ�ȷ�����������˵�ľ���ؽ�������������Ϊ���һ��Ӧ���о��ṩһ�����������ݡ�
1 ���Ϻͷ���
1.1 ����
1.1.1 ����
�����õ�3��ϸ��Ϊ��������ǰ�ڴ��������(��ɳ��빲���ݱ���)ʴ�������з���ɸѡ�õ��ľ����ֱ�Ϊ�����˾�������B-2(Acinetobacter sp. B-2)�������Ͼ�������B-6(Pandoraea sp. B-6)�����ʰ����˾�������B-7(Novosphingobium sp. B-7)��
1.1.2 ������
(1) ���ֱ�����������ľ���ػ�����(����й⸴��ϸ�����о�������)3 g/L��K2HPO4 1 g/L��MgSO4��7H2O 0.2 g/L��CaCl2 0.1 g/L��FeSO4��7H2O 0.05 g/L��MnSO4 ��H2O 0.02 g/L��KH2PO4 1 g/L��(NH4)2SO4 1.98 g/L����֬15 g/L��pHΪ7.0���ҡ�
(2) ���ֻ��������LB������(������10 g/L����ĸ����5 g/L��NaCl 10 g/L��pHΪ7.0����)��
(3) Һ�彵����������ľ���ػ�����3 g/L��K2HPO4 1 g/L��MgSO4��7H2O 0.2 g/L��CaCl2 0.1 g/L��FeSO4��7H2O 0.05 g/L��MnSO4 ��H2O 0.02 g/L��KH2PO4 1 g/L��(NH4)2SO4 1.98 g/L��pHo 7.0���ҡ�
(4) ����������(BM������)����ĸ��10 g/L��������20 g/L����֬15 g/L��pHΪ7.0���ҡ�
1.2 ���鷽��
1.2.1 ľ���ؽ���ø�Ķ��Լ��
���ñ�����(Azure-B)�� RB����(Remazol Brilliant Blue)Ⱦ��ƽ���⣬��BM�������зֱ����0.1 g/L��Ⱦ�ϱ�������RB��������ƽ�����ָ����꣬��30 ���������о���������ÿ��۲죬��ƽ���������о�����Χ��ɫȦ�����������Լ��ľ���ؽ���ø�Ƿ����������ɫȦ������Ϊ(+)������ɫȦ������Ϊ(-)��
1.2.2 �����ľ���ؽ�������
��LB�����������ϸ���������ڣ����������1%������Һ�彵���������У���30 ��ҡ����������ÿ��ȡ���ⶨ����Һ��ľ���ػ����Ƶ�Ũ�ȣ�ֱ��ľ���ػ�����Ũ�Ȳ��ٱ仯Ϊֹ��
1.2.3 ľ���ؽ���ĵ����о�
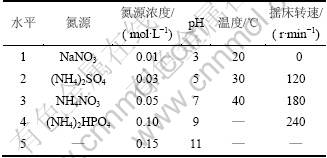
��LB�����������ϸ���������ڣ����������Ϊ1%�����ڽ�������Һ�����������и����������ѡȡ��Դ����ԴŨ�ȡ�pH���¶ȡ�ҡ��ת��5�����ؽ���ϸ��ľ���ؽ���ʵ�顣��������ѡȡˮƽ����1��
��1 �����������ؼ���ѡˮƽ
Table 1 Incubating control factors and parameter levels
1.3 �ⶨ����
1.3.1 ľ���ػ�����-��̼�����߲ⶨ
ȷ��ȡľ���ػ�����1.000 0 g��������ˮ������1 L�������ʵ�ϡ�ͣ��õ�ľ���ػ���������Ũ��Ϊ0.1~0.8 g/L�ĵ��ݶ���Һ���ֱ�ⶨ����Һ����̼(Total carbon��TC)����Ũ�ȡ���ľ���ػ���������Ũ��Ϊ�����꣬��̼����Ũ��Ϊ�����꣬����ľ���ػ�����-��̼����Ũ�ȱ����ߡ�
1.3.2 ľ���ؽ����ʲⶨ
ȡ����Һ4 mL����ת��12 000 r/min����6 min������Һ��ֱ��Ϊ0.22 ��m������ͷ(�Ϻ��������﹤�̼�����������˾)���ˣ��ⶨ��ҺTC����Ũ�ȡ�����ľ���ػ�����-��̼����Ũ�ȱ����ߣ������ľ���ػ����Ƶ�����Ũ�ȡ����ݽ���ǰ��ľ���ػ���������Ũ�ȵıȽϣ����ɵõ�ľ���ؽ����ʡ�������Ҫ�����ÿ�����ͬʱ����3��ƽ��ʵ�飬���ȡƽ��ֵ��
1.3.3 ��Һ��̼�ⶨ
��Һ��̼����Ũ�Ȳ���TOC��VCPH�����л�̼������(�ձ�����˾����)�ⶨ��
2 ���������
2.1 ľ���ؽ���ø�Ķ��Լ����
ľ���صĽ�����Ҫ������һϵ��ø�Ĺ�ͬ���ã���Щø��Ҫ����ľ���ع�������ø(Lignin peroxidases, LiP)���̹�������ø(Manganese peroxidases, MnP)����ø(Laccase, Lac)��ͨ�����Լ������ľ���ؽ���ø����������Գ����жϾ����ľ���ؽ�������������������ɫ��LiP��MnP�IJ����йأ�������ӳ��ø�IJ�������ø��ʹRB������ɫ[11-13]��������2��ʾ������Acinetobacter sp. B-2��Pandoraea sp. B-6�Ա������н�ǿ����ɫ��������RB��������ɫ���á�Novosphingobium sp. B-7�Ա�������RB��������һ������ɫ����������ƽ�����ɫ�������������ľ�����յ�����ڵ������3��������в�ľ���ؽ���ø�����������ݱ�����ƽ����ɫ��Ӧ��RB������ɫ��Ӧֻ�ܼ��жϾ���IJ�ø�����Ҫ����ȷ�������ľ���ؽ��������������ľ���ؽ���Ķ����ⶨʵ�顣
��2 ����ƽ������Ⱦ�ϵ���ɫЧ��
Table 2 Effects of bacteria strains on decolorization of dye under plate culture

2.2 ľ���ػ�����-��̼����Ũ�ȱ�����
ľ���ػ���������̼����Ũ�Ȳⶨ�����ͼ1����ͼ1�ɼ���ľ���ػ�������Һ����Ũ����TC����Ũ���кܺõ�������ԣ���������Ʒľ���ػ���������Ũ�ȵ�ȷ����
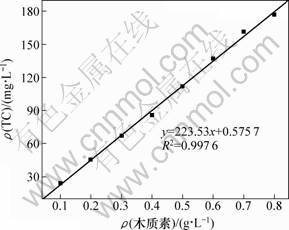
ͼ1 ľ���ػ�����-��̼����Ũ�ȱ�����
Fig.1 Standard curve of mass concentration of sodium lignin sulfonate-total carbon
2.3 ����ľ���صĽ�������
3��ϸ�����ֵ�Һ�彵����������ÿ��ȡ���ⶨ��ľ��������Ũ�ȣ�3�����ľ���ؽ��������ͼ2��ʾ����ͼ2�ɼ���������ǰ3��Ϊ3�����ľ���ؿ��ٽ����ڣ���4�������ľ���ؽ����������ڻ��ͣ�����ľ���ص�����Ũ�Ȼ������ٱ仯�������ľ���صĽ�������ȶ����ɼ���3��ϸ����ľ���صĽ�����Ҫ�����ڳ�����л�Σ�����Ramachandra��[14]��Ϊϸ����ľ���ؽ��ⷢ���ڳ�����л�ε��о�������һ�¡���4���Ժ�����ϸ����������˥���ڣ�����ϸ����ʼ�������������ܣ��ᵼ����Һ��TC����Ũ�����ߣ�Ӱ��ʵ�����IJⶨ������ʵ���������ľ���ؽ����ʣ���ˣ�ѡ������3 d��ⶨ�������¾����ľ���ؽ����ʡ�Ŀǰ���������й�ľ�������オ����о���༯���������������������ڳ������������Ҫ����3���������ò��ܼ�ľ���صIJ��ֽ���[15-16]��ϰ��÷��[17]��ũ�ַ����з���õ�1�����ù���ڹ�̬��������������30 d��ľ���ؽ����ʴ�16.87%��ͨ����Ϊ�Ƿ�˿״ϸ������ľ���صĵ���Է���������ֺ�ľ���صĽ����м����[18]�����ȷ��[19]����ֽ��Һ��ɸѡ����õ�6��ϸ��Һ�壬����12 d��ľ���صĽ�����Ϊ10%~20%�����о����õĴ��������ʴ�������з���ɸѡ�õ���3���˿״ϸ����5���ľ���صĽ�����������ȶ������������Ⱦ��н������ʿ졢����ʱ��̵��ص㡣Ŀǰ����������ڷ�˿״ϸ����ľ���ؽ����о����٣�������һ�������ľ���ؽ�����������������Լ�����ľ���ص���Ȼ��������������û�ȱ���˽⣬��ˣ����ھ��и�Чľ���ؽ��������ķ�˿״ϸ�����о�������Ҫ�����塣
2.4 ��Դ��3���ľ���ؽ����Ӱ��
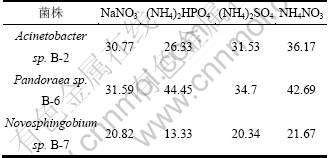
��ʵ���е�3��ϸ������������ľ���ػ�����ΪΨһ̼Դ����ѡ�����������Դ���е�Դѡ��ʵ�顣�ֱ��������ơ��������李�����李������(��Ϊ0.03 mol/L)��ΪΨһ��Դ���ⶨ3�����ľ���ؽ����ʣ�ʵ��������3���ɼ��������Ϊ����Acinetobacter sp. B-2��Novosphingobium sp. B-7����ѵ�Դ����������Ϊ����Pandoraea sp. B-6��ѵ�Դ�����Ե�ԴΪ�����ʱ��3�����ľ���ؽ����ʶ���Խϸߡ������ӵ�[20]��1���ȼ�ľ���ؽ������ø�����ľ���ؽ�����о�Ҳ���������Ϊ�þ�����Ѳ�ø��Դ������Acinetobacter sp. B-2��Pandoraea sp.B-6��Ȼ��4�ֲ�ͬ��Դ������ľ���ؽ����������Բ�𣬵����彵���ʶ��ϸߣ�������2�����Դ���з�רһ�ԡ�
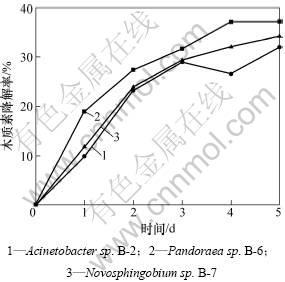
ͼ2 ����ľ���ؽ�������
Fig.2 Lignin-degrading curves of bacteria strains
��3 ��Դ�Ծ���ľ���ؽ����ʵ�Ӱ��
Table 3 Effect of nitrogen source on lignin degradation by bacteria strains %
2.5 ��ԴŨ�ȶ�3���ľ���ؽ����Ӱ��
��ľ���ػ�����Ũ�Ȳ���������£�ͨ���ı���ѡ����3��ϸ�����Ե���ѵ�Դ(Acinetobacter sp. B-2��Novosphingobium sp. B-7(̼ԴΪ�����)��Pandoraea sp. B-6(̼ԴΪ��������)��Ũ�������ڽ����������е�̼���ȡ�ѡȡ�ӽϵ�Ũ�ȵ���Ũ��5���ݶȣ��ֱ�ⶨ3�����ľ���ؽ��������ʵ������ͼ3����ͼ3�ɼ�����ͬ�ĵ�ԴŨ�ȶ���ľ���ؽ�����һ����Ӱ�죬������ѡ��ԴŨ�ȷ�Χ�ڲ�δ���ֳ�ľ���ؽ����ʺ͵�ԴŨ�ȵ�����ԡ��ܵ���˵���͵�Ũ���ƺ������ھ���ľ���ؽ�����������ߡ���Acinetobacter sp. B-2��Novosphingobium sp. B-7����ԴŨ��ѡ��Ϊ0.01 mol/L����Pandoraea sp. B-6����ԴŨ��ѡ��Ϊ0.03 mol/L���ܶ������漰����ԴŨ�ȶԾ����øӰ����о���һ����Ϊ�����ľ���ؽ��ⷢ���ڴμ���л�Σ���̼����������ľ���صĽ�����ø�����ε�[21]�о���Ӫ����������No.4510��LiPøӰ�죬���������̼������LiPø�IJ����������ߵ�[22]�Ի���ԭëƽ�����ľ���ؽ���ø���о����֣������ڵ����������¶����ڽϸߵ�Ũ����������������¶�������LiPø���ԡ��ɼ�����ͬ������ľ���ؽ������ܼ�ľ���ؽ���ø�IJ������ܴ�
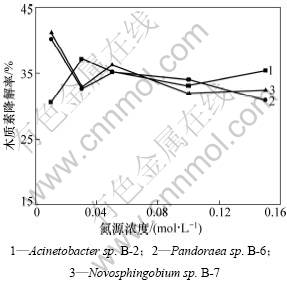
ͼ3 ��ԴŨ�ȶ�3���ľ���ؽ����Ӱ��
Fig.3 Effect of nitrogen source concentration on lignin degradation by bacteria strains
2.6 �¶ȶ�3���ľ���ؽ����Ӱ��
����Ŀǰ����3��ϸ��ľ���ؽ�����о��������٣���ľ���صĽ��������в��������ˣ�����ɸѡ�������ʱ�Ļ�������������̽����20��30��40 �� 3���¶��¾����ľ���ؽ������ܣ�ʵ������ͼ4��ʾ����ͼ4���Կ������¶ȶԾ���ľ���ؽ������ܵ�Ӱ��ܴ��Ҷ�3�����Ӱ�����ƻ�����ͬ����30 ��ʱ����Ч�����Ժ���20 ���40 ��Ľ���Ч����������3����������¶ȷ�Χ�Ͽ������¶�Ϊ20 ���40 ��ʱ��3 d����20%���ҵĽ����ʡ�
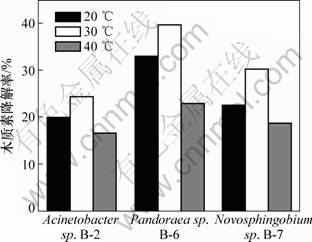
ͼ4 �¶ȶ�3���ľ���ؽ����Ӱ��
Fig.4 Effect of temperature on lignin degradation by bacteria strains
2.7 ��ʼpH��3���ľ���ؽ����Ӱ��
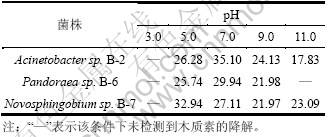
�����Ե����Ե���Һ�彵���������ij�ʼpH(3.0��5.0��7.0��9.0��11.0)�������������ֲ��䣬�ֱ�ⶨ3�����ľ���ؽ����ʣ�ʵ��������4���ӱ�4���Կ���������ʼpHΪ3ʱ��3�����δ��ʾľ���صĽ������ܣ�����Acinetobacter sp. B-2��Pandoraea sp. B-6�ڳ�ʼpHΪ7ʱ������Ч����ã���Novosphingobium sp.B-7�����ڳ�ʼpHΪ5ʱ����������ߣ�����Acinetobacter sp.B-2��Novosphingobium sp.B-7�ڳ�ʼpHΪ11ʱ���Կ��Լ���20%����ľ���ر����⣬������Pandoraea sp. B-6���ܼ�ľ���ؽ���Ч������ֽ��ˮһ��ʼ��ԣ��������Щ�������Զ��ڹ�ҵӦ�ü������������⣬ͨ���ⶨ3��������������������pH(��5)���֣����ܳ�ʼpH��ƫ���Ի�ƫ���ԣ���ľ���ؽ��������Һ����pH����תΪ���Ե����ơ������϶���������ľ���ؽ�����������Զ�����pH�Ĺ��ܣ��Ա�֤������Ч�ط���ľ���ؽ���������
��4 ��ͬ��ʼpHʱ�ľ���ľ���ؽ�����
Table 4 Lignin degradation rate by bacteria strains at different initial pH values %
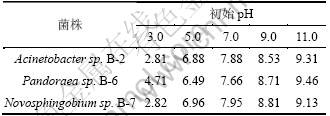
��5 ����ľ���ؽ���������pH
Table 5 Final pH after lignin degradation by bacteria strains
2.8 ҡ��ת�ٶ�3���ľ���ؽ����Ӱ��
�ı�ҡ��ת�٣���������Ӫ�����ʵĴ��ʷ��濼����������������������ԡ��ֱ���60��120��180��240 r/min 4��ת�����������겢�����ľ���ؽ������ܡ���ͼ5�ɼ���3���ҡ��������ľ���ؽ���Ч�����Ժ��ھ�ֹ����ʱ�Ľ���Ч��������������ʱ��ת��Ϊ120 r/minʱľ���صĽ����Ѿ����͡����������Ӱ���˾��������������ʹ������ľ���ؽ��ܽ�ϱ�����ѣ��Ӷ��谭�˾����ľ���صĽ��⡣120 r/min��ҡ��ת�ٿ�����Ч�شٽ����������������������Ӫ�����ʵĴ��ݣ���3�־���������ľ���ؽ������ʵ��ġ�
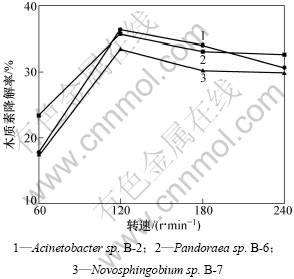
ͼ5 ҡ��ת�ٶԽ���ľ���ص�Ӱ��
Fig.5 Effects of rotation speed on lignin degradation by bacteria strains
3 ����
(1) Acinetobacter sp. B-2��Pandoraea sp. B-6��Novosphingobium sp. B-7��ʹ��������RB����ƽ����ɫ������ľ�����յ��������¾��в�ľ���ؽ���ø��������
(2) ��5��ʱ��3��ϸ����ľ���صĽ�����������ȶ������нϿ�Ľ����ٶȣ�����Ч�ʱ�Ŀǰ�����Ķ������ֵĽ���Ч���á�
(3) ���õ�����ʵ���3��ϸ��������ľ���ؽ��������ĵ����о�������ȷ����3������˵Ľ�����������Acinetobacter sp. B-2����ԴΪ����泥���ԴŨ��Ϊ0.01 mol/L�������¶�Ϊ30 �棬��ʼpH=7.0��ҡ��ת��Ϊ120 r/min����Pandoraea sp. B-6����ԴΪ�������泥���ԴŨ��Ϊ0.03 mol/L�������¶�Ϊ30 �棬��ʼpH=7.0��ҡ��ת��Ϊ120 r/min����Novosphingobium sp. B-7����ԴΪ����泥���ԴŨ��Ϊ0.01 mol/L�������¶�Ϊ30 �棬��ʼpHΪ5.0��ҡ��ת��Ϊ120 r/min�������˵Ľ��������£�3�����3�콵���ʾ��ɴﵽ30%~35%�����н�ǿ��ľ���ؽ���������
�ο����ף�
[1] Sjostrom E. Wood chemistry, fundamentals and application[M]. New York/London: Academic Press, 1993: 107-121.
[2] Kirk T K, Schultz E, Connors W J, et al. Influence of culture parameters on lignin metabolism by Phanerochaete chrysosporium[J]. Archieve of Microbiology, 1978, 117: 277-285.
[3] �����, ���滪. ����� F2����������ľ���ؽ���ø���Ե��о�[J]. ������ѧѧ��, 2005, 25(2): 226-231.
LI Cui-zhen, WEN Xiang-hua. Characterization of growth and ligninolytic enzymes production of a white rot fungus F2[J]. Acta Scientiae Circumstantiae, 2005, 25(2): 226-231
[4] �����. ���������ѧ�����\��[M]. ����: ��ѧ��ҵ������, 2005: 10-30.
LI Hui-rong. Biology and biotechnology of white rot fungi[M]. Beijing: Chemical Industry Press, 2005: 10-30.
[5] �Ƶ���, ������, �ƹ���, ��. ������̬�����������Ż����併��ֲ�������ʵ��о�[J]. ������ѧѧ��, 2005, 25(2): 232-237.
HUANG Dan-lian, ZENG Guang-ming, HUANG Guo-he, et al. Optimum conditions of solid-state fermentation for white-rot fungi and for It��s degrading straw[J]. Acta Scientiae Circumstantiae, 2005, 25(2): 232-237.
[6] ��Ծ��. �������ʴ������ķ����������ľ���ؽ��������о�[D]. ��ɳ�����ϴ�ѧұ���ѧ�빤��ѧԺ, 2009: 13-15.
CHEN Yue-hui. Identification of microorganisms isolated from erosive bamboo slips Kingdom Wu and characterization of lignin degradation[D]. Changsha: Central South University. School of Metallurgy Science and Technology, 2009: 13-15.
[7] Delneri D, Degrassi G, Rizzo R, et al. Degradation of trans-ferulic and p-coumaric acid by Acinetobacter calcoaceticus DSM 586[J]. Biochimica et Biophysica Acta, 1995, 1244: 363-367.
[8] Vasudevan N, Mahadevan A. Degradation of lignin and lignin derivatives by Acinetobacter sp.[J]. Journal of Applied Bacteriology, 1991, 70: 169-176.
[9] Siddique T, Benedict C O, Arshad M. Biodegradation kinetics of endosulfan by fusarium ventricosum and a Pandoraea Species[J]. Agric Food Chem, 2003, 51(27): 8015-8019.
[10] Stolz A. Molecular characteristics of xenobiotic-degrading sphingomonads[J]. Appl Microbiol Biotechnol, 2009(81): 793-811.
[11] ����, ������, ����Ƽ, ��. ���ּ���ľ���ؽ����������ɸѡ����[J]. ����ѧͨ��, 2002, 29(1): 67-69.
CAI Lei, YIN Jun-feng, YANG Li-ping, et al. Several qualitative methods for the screening of fungi to decompose lignin[J]. Microbiology China, 2002, 29(1): 67-69.
[12] Archibald F S. A new assay for lignin-type peroxidases employing the dye Azure B[J]. Appl Environ Microbiol, 1992, 58(9): 3110-3116.
[13] ���ӱ�, ������, ������, ��. һ�������������ø��RB��������ɫ����[J]. Ӧ���뻷������ѧ��, 2002, 8(3): 298-301.
KANG Cong-bao, LI Qing-xin, LIU Rui-tian, et al. Decolorization of remazol brilliant blue R by laccase produced from a white rot fugus[J]. Chin J Appl Environ Biol, 2002, 8(3): 298-301.
[14] Ramachandra M, Crawford D L. Hertel G. Characterization of an extracellular lignin peroxidase of the lignocellulolytic actinomycete Streptomyces viridosporus[J]. Applied and Environmental Microbiology, 1998(54): 3057-3063.
[15] Ferraz A, Duran N. Lignin degradation during softwood decaying by the ascomycete Chrysonilia Sitophila[J]. Biodegradation, 1995, 6(4): 265-274.
[16] Dey S, Malti T K, Bhattacharyya B C. Production of some extracellular enzymes by a lignin peroxidase-producing brown-rot fungus and its comparative abilities for lignin degradation and dye decolorization[J]. Applied and Environmental Microbiology, 1994, 60(11): 4216-4218.
[17] ϰ��÷, ������, ������, ��. ����ùAspergillus nigerľ����ά�ؽ�����������ø�о�[J]. ũҵ������ѧѧ��, 2007, 26(4): 1506-1511.
XI Xing-mei, ZENG Guang-ming, YU Hong-yan, et al. Lignocelluloses degrading ability of Aspergillus niger and the enzyme production[J]. Journal of Agro-Environment Science, 2007, 26(4): 1506-1511.
[18] Toumela M, Vikman M, Hatakka A, et al. Biodegradation of lignin in a compost environment: A review[J]. Bioresource Technology, 2000, 72: 169-183.
[19] ���ȷ�, ��־��, ���. ��ֽ��Һľ���ؽ�������ķ���ͽ��������о�[J]. ��������, 2002, 20(3): 78-80.
SUN Xian-feng, ZHANG Zhi-jie, CUI Hong-jun. Study on isolation of lignin-degradation microorganisms of pulping liquor and degradation characteristics[J]. Environmental Engineering, 2002, 20(3): 78-80.
[20] ������, �ż�ҫ, ������, ��. �ȼ�ϸ������ľ���صĸ���̼Դ����л�о���: ����̼Դ��Ϸ�ʽ����Դ��ѡ��[J]. ���л����������̬, 2000, 13(2): 8-10.
LUO Yu-xuan, ZHANG Jia-yao, GUAN Xiao-wu, et al. Research of compounded carbons cometabolism of alkaliphilic ligninolytic bacteria ��: Selections of compounded carbons and nitrogen sources[J]. Urban Environment and Urban Ecology, 2000, 13(2): 8-10.
[21] ����, ·��ƽ, ������. ����ľ�ع�������ø������������øҺ�Ե��ݽ�����о�[J]. ��ά�ؿ�ѧ�뼼��, 2002, 10(4): 41-48.
BI Xin, LU Fu-ping, Du Lian-xiang. Optimum conditions for production of lignin peroxidase and degrading rice straw with crude enzyme of LiP[J]. Journal of Cellulose Science and Technology, 2002, 10(4): 41-48.
[22] ������, ���滪, ���, ��. ����ԭëƽ����ڶ��ְ���Ũ����ľ���ؽ���ø�IJ���[J]. ������ѧѧ��, 2003, 23(6): 802-806.
YU Guo-ce, WEN Xiang-hua, LI Dong-feng, et al. Formation of the ligninolytic enzymes by Phanerochaete chrysosporium under various ammonium nitrogen concentrations[J]. Acta Scientiae Circumstantiae, 2003, 23(6): 802-806.
(�༭ �²ӻ�)
�ո����ڣ�2010-11-15�������ڣ�2011-03-01
������Ŀ�����Ҹ����о���չ�ƻ�(��863���ƻ�)�ص���Ŀ(2007AA021304)������ʡ�Ƽ��ƻ��ص���Ŀ(2009WK2010)������ˮ����Ⱦ�����������Ƽ��ش�ר��(2009ZX07212-001-01)
ͨ�����ߣ�����Ԫ(1966-)���У����������ˣ����ڣ���ʿ����ʦ�����·�ˮ���������о����绰: 0731-88836921��E-mail��lychai@csu.edu.cn
ժҪ�����ñ�������RB����ƽ��Դ��������ʴ���з���õ���3��ϸ��Acinetobacter sp. B-2��Pandoraea sp. B-6��Novosphingobium sp. B-7������ɫ���飬������3��ϸ����Һ�����������µ�ľ���ؽ������ܣ�������ľ���ؽ��������ĵ��ؽ����о�������ȷ����3������˵Ľ����������о������������3��ϸ������ʹ��������RB����Ⱦ����ɫ���������ܹ���ľ���ؽ���ø����3��ϸ����ľ���ؽ������ʽϿ죬��5��ľ���صĽ�����������ȶ����������˵Ľ����������£���Acinetobacter sp. B-2����ԴΪ����泥���ԴŨ��Ϊ0.01 mol/L�������¶�Ϊ30 �棬��ʼpH=7.0��ҡ��ת��Ϊ120 r/min����Pandoraea sp. B-6����ԴΪ�������泥���ԴŨ��Ϊ0.03 mol/L�������¶�Ϊ30 �棬��ʼpH=7.0��ҡ��ת��Ϊ120 r/min����Novosphingobium sp. B-7����ԴΪ����泥���ԴŨ��Ϊ0.01 mol/L�������¶�Ϊ30 �棬��ʼpHΪ5.0��ҡ��ת��Ϊ120 r/min�������˵Ľ��������£���3��ϸ����3���ľ���ؽ����ʾ��ɴﵽ30%~35%��
