һ���ͭ��ר������ϸ���ķ��������
�� ��1, 2��������1���� ��1�������1, 2���ƾշ�2��������1, 2
(1. ���ϴ�ѧ ��Դ�ӹ������﹤��ѧԺ������ ��ɳ��410083��
2. ���ϴ�ѧ ����ұ��������ص�ʵ���ң����� ��ɳ��410083)
ժ Ҫ���Ӹ�������������ˮ�з���õ�һ���ܹ������ϸ��(����ΪBY)���þ���Ϊ����������ϸ�����̸�״�������ֱ��������Ϊ(0.45��0.05) ��m��(1.5��0.4) ��m, ���������¶�Ϊ25~30 �棬����pHֵΪ2.0������ �����ͣ�������������������͵�Ũ�ȵ�����������������������������ơ��������������о����������BY��������������������˾�(���A. f��)WJ13λ��ϵͳ������ͬһ����֧�У����ƶ�Ϊ99.66%�����A. f����ATCC23270�����ƶ�Ϊ99.93%���������ĺ��������뱨������(�Ǽ����к�DQ166839-DQ166841)����1�������������������ȫһ�£���30 �����27 d�������ҡƿ��Cu2+������Ũ�ȼ��ﵽ60 mg/L����������ʱCu2+������Ũ�Ƚ�Ϊ10 mg/L�������þ������Ӧ���ڻ�ͭ�������DZ����
�ؼ��ʣ���������������˾��������������������ø����16S rRNA���У���ͭ��
��ͼ����ţ�Q939 ���ױ�ʶ�룺A ���±�ţ�1672-7207(2007)04-0639-06
Isolation and characterization of bacterium for chalcopyrite bioleaching
YANG Yu1, 2, WAN Min-xi1, PENG Hong1, QIU Guan-zhou1, 2, HUANG Ju-fang2, HU Yue-hua1, 2
(1. School of Minerals Processing and Bioengineering, Central South University, Changsha 410083, China;
2. Key Laboratory of Biometallurgy of Ministry of Education, Central South University, Changsha 410083, China)
Abstract: A bioleaching bacterium was isolated from a sulfide tailings of Baiyin, Ganshu Province in China, named as BY. The strain BY is gram-negative and rod-shaped in (0.45��0.05) ��m��(1.5��0.4) ��m. The optimum growth temperature is 25-30 ��, and the optimum growth pH is 2.0. It can grow autotrophically by using ferrous, sulfur and glucose as sole energy sources, but it can not grow with NaS2O3 and peptone. In the phylogenetic tree, BY has similarity to Acidithiobacillus ferrooxidans (A.f) strain WJ12, and the type strain ATCC23270 with 99.93% sequence similarity. There is only one in coding sequence (CDS) and the same protein sequence. The copper concentration is 60 mg/L while the control is only 10 mg/L after 27 d. This bacteria can be used to leach chalcopyrite in industry application.
Key words: Acidithiobacillus ferrooxidans; bioleaching; Iro protein gene; 16S rRNA sequences; chalcopyrite
��ͳ�Ļ���ͭ�������ڶԻ��������Ⱦ�������Ͼ�����ս����������ʪ����ͭ���о��ܵ��ձ��ע����ͭ������Ȼ�糣����ͭ����֮һ��������������ṹ���������ʺ������ʻ�ͭ������ѳ�Ϊͭ��ҵ�о����ȵ㣬Ŀǰ����ͭ��ʪ��������������������������Ȼ����������������Լ���ԭ������[1-2]���������������������Ⱦ������̶̡����ռ��ײ�����Ͷ���١��ܺ�С���ɱ��ͺͶԻ����Ѻõ��ŵ㣬���ҹ�ͭ����Դ���Ʒλ��[3]���ʺϲ��������������������Ȼ������ͭ����Ŀǰ�Ƚ��ѽ����Ŀ���[2, 4]�����ڻ�ͭ���������������ص㣬ѡ�������Խ�����ͭ��ĸ�Чר�������Ƿdz���Ҫ�ġ�
��������������˾�(Acidithiobacillus ferrooxidans�����A. f��)��������ԭ������硢����Ӫ��ԭ�������š�ϸ���١���ϸ���ơ���˾�������Ҫ��л�ص��ǣ�̼ԴΪCO2����ԴΪNH+4��������Fe2+��S0��S2���Ļ�����Ȼ��������������[5]����1947����Colmer��Hinkle���ȷ��봿�������������� A. f��һֱ�����������������ú����������Ҫ��ϸ��֮һ[6-8]��
�о�����������ͬ���������£���ͬ��Դ����������������˾�����Խ�������Ľ���Ч���������Բ���[5-6]����ˣ��ɼ���ͬ�����ˮ����ɸѡ����Ӧ�ڲ�ͬ�������õ��������꣬���Բ�ͬ����������������ͬ����ͻ�����A.f���Ķ�����(������Ⱥ�����ԡ�ϵͳ���������Ժ��ܶ�����)����̬�ݻ����ɿ�չ�о������ڽ�һ���������������Ⱥ��ṹ���ܡ�������Ч������������Ҫ����[9]���ڴˣ��������ߴӸ������������ij����ˮ���з��봿����һ����������ϸ���������������������������о���ͬʱ�������Ի�ͭ��Ľ���������
1 ���Ϻͷ���
1.1 ���ַ���
����Ӹ�����������Ŀ��ˮ�з���õ�������ΪBY���ÿ����п�ʯΪ�����������ʯ����Ҫ�����У���ͭ��������ʯ�����ˮ�¶�ԼΪ25 �棬pHֵΪ6.6���������ԡ��������Ҫ�����ǣ����ɼ���������ˮ������9KҺ��������[10]��������������������pHֵ�½���1.0���Һ����ݶ�ϡ�ͷ��ڸĽ���9K����������Ϳ��[11]������ƽ�廮�߷����롣
1.2 ������
����Һ��������(9K)���䷽���£�(NH4)2SO4����Ũ��Ϊ3 g/L��KCl����Ũ��Ϊ0.1 g/L��K2HPO4����Ũ��Ϊ0.5 g/L��MgSO4?7H2O����Ũ��Ϊ0.5 g/L��Ca(NO3)2����Ũ��Ϊ0.01 g/L��FeSO4?7H2O����Ũ��Ϊ44.7 g/L��pHֵΪ1.8~2.0��
����������Ϊ�Ľ���9K����������������ÿ��9KҺ���������м���15 g��֬��15.4 g KSCN��pHֵΪ2.0~2.2������������Ϊ����FeSO4��9K��������
1.3 ������������
a. �����¶ȡ�����ͬ�ľ������ֵ�9KҺ���������У��ֱ����ڲ�ͬ�¶��£�����3 d����Ѫ�������������ⶨ��ͬ�¶���ϸ��������״����
b. ���ʳ�ʼpHֵ����1 mol/L���Ὣ9KҺ��������������ͬpHֵ��������ͬ�ľ���������3 d����Ѫ�������������ⶨ��ͬpH�¾�������״����
c. �����������ԡ�����������(����Fe2+��9K������)�зֱ���뵰����(0.1%)��������(0.1%)�����(5%)��NaS2O3(1%)��FeSO4?7H2O(14.7%)��FeSO4?7H2O(14.7%)+������(0.1%)��FeSO4?7H2O (14.7%)+������(0.1%)�����У�NaS2O3��FeSO4?7H2O���˳������������������Ǹ�ˮ����1 h������ټ�������������BY������Һ���֣�ÿ����3 d����1�Σ���������3�Σ�ͿƬ���������¹۲�ϸ�������������
1.4 �羵�۲�
��30 ��Һ��������ϸ�������������ڣ��ռ����壬��������Ⱦɫ���ڹ�ѧ�����¹۲졣������Ϳ����С��Dz�Ƭ�ϣ���Ȼ���ɺ����õ��罺����Բ����Ƭ�ϣ�������ɨ��羵�¹۲졣
1.5 ���������������ԵIJⶨ
1.5.1 �����������Բⶨ
ȡBY�����Һ���������5��10-4 mol/L K2Cr2O7�����е�Fe2+�������еζ���ÿ��һ��ʱ��(һ��Ϊ3~4 h)ȡ0.5 mL��Һ��������ˮ(����1%ϡ����)ϡ��10������ȡ���е�1 mL�ζ���ÿ�εζ��ظ�3�Σ�Ȼ��ȡ��ƽ��ֵ��Ϊ�ζ�ֵ���ζ���1?1��������Ϊ��ϼ�����0.2%��������������ָʾ�����ζ�������ɫ�����ζ��յ㡣
1.5.2 ���������Բⶨ
��BY������ֵ�ֻ���������9K����������(�����������Ϊ5%)�У�ͨ��pHֵ�ı仯������BY��������������ԡ�
1.6 ��������
����Ϊ��ͭ�����ȷֲ�Ϊ104~147 ��m��������Ϊ10%�����ڳ�ʼ��Ũ��Ϊ1.0��107 ��/mL�������������£�250 mLҡƿװҺ100 mL��������Ϊ5%��ת��Ϊ160 r/min���¶�Ϊ30 �档
1.7 ϸ����������ȡ��PCR����
1.7.1 ������DNA��ȡ
�Դ����������ľ���ʹ��Sangon��˾��UNIQ-10��ʽ������DNA�����Լ�����ȡ������DNA������1%������Ӿ���������
1.7.2 16S rRNA����
������ϵ(50 ��L)Ϊ��33 ��L��ˮ��10��PCR buffer 5 ��L��10��dNTPs 5 ��L������FC27��RC1492��2 ��L(5 pmol/��L)��Taq DNA�ۺ�ø 1 ��L��BY����Ļ�����DNA 2 ��L����������Ϊ��94 �棬5 min��94 �棬1 min��55 �棬1 min��72 �棬3 min(��30��ѭ��)��72 �棬10 min��PCR����Ϊ16S rRNA��������ͨ������[8]��
��������(FC27)��5��-AGAGTTTGATCCTGG-��CTCAG-3����
��������(RC1492)��5��-TACGGCTACCTTGT-��TACGACTT-3����
1.7.3 ��������ø��������
������ϵ(50 ��L)Ϊ��33 ��L��ˮ��10��PCR buffer 5 ��L��10��dNTPs 5 ��L������YT1��YT2������2 ��L(5 pmol/��L)��Taq DNA�ۺ�ø 1 ��L��BY����Ļ�����DNA 2 ��L����������Ϊ��94 �棬2 min��94 �棬40 s��59 �棬30 s��72 �棬40 s(��35��ѭ��)��72 �棬5 min��������Ʋο�NCBI�б�����(������ΪEO3451����СΪ500 bp)������Primer5������ơ�
��������(YT1)��5��-CTCTGACCGGCGAATC-��GGG-3����
��������(YT2)��5��-CCAACCGCATCCGCAT-��ATCTTG-3����
1.8 ���������
����PCR�����¡�Լ���(�Ϻ�����SK2212)�������õ���16S rRNA����������ø����Ƭ�ν��п�¡������ΪpUCm-T(PCR�����¡�Լ����Դ�)������ɸѡ�����õ������¡���ɱ�������Զ־���\���������ι�˾����16S rRNA�����Ѿ��ύ��GenBank(��¼��ΪDQ062114)��������16S rRNA������GenBank�����������ݿ�������бȽϣ���clustalx1.8��������ȫ���бȶ�[12]��������ϵͳ����������������ø���������Ѿ��ύ��GenBank(��¼��ΪAY864809)��������������GenBank�����������ݿ�������бȶԡ�
2 ���������
2.1 ������̬��������������
�����Ǹ���������ϸ�����̸�״�������ֱ��������Ϊ(0.4��0.1) ��m��(1.4��0.2) ��m���ڸĽ���9K�����������������������Բ�Σ�ֱ��Ϊ0.6~1.0 mm����Ե�������������dz��ɫ(��ͼ1��ʾ)�����������¶�Ϊ25~30 �棬����pHֵΪ2.0��������������������������������������������ǻ����������Ҳ���������������õ������������������������˻����������Ҳ��������(���1��ʾ)�����������Ƕ�BY����������Ӱ�죬�ʵ��Ӵ�����������+�����ǻ���������������ǵĺ�������������Ũ��Ϊ3%ʱ�����������Ѽ��٣����������������Ƕ�BY������������������á�

ͼ1 BY����ľ�����̬(a)��ɨ��羵ͼ��(b)
Fig.1 Colony of BY(a) and scanning electron
micrograph image(b)
��1 BY������Դ��������
Table 1 Characteristics of BY strain using energy material
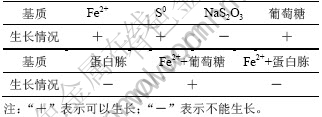
2.2 �����������Ժ�����������
2.2.1 ������������
BY������н�ǿ�������������ԣ��������������������������ء�ͼ2��ͼ3��ʾ�ֱ�Ϊ��30 �棬pHֵΪ1.8��9KҺ���������У�BY������������ߺ��������������ߡ���Ϸ���ͼ2��ͼ3���֣�7 hʱ��ʵ����Ϳհ����������Բ����������ϸ����ת�Ƶ��»�����, ������ֳ���ڳ����ڣ���7~50 h֮�䣬����ʼ��������������ʱ������ӣ����������ʻ�����ߣ���հ��յIJ��������ֳ�������50 hʱ������������ڣ�����������Ҳ�������ߣ�70 h�Ժ����������ʴﵽ99.5%��ͼ3��ʾΪBY������30 �棬pHֵΪ2.0��9K�������е���������������ͼ��ͼ��BY�����ǽ�����BY�������������Fe2+��������ʱ��ı仯���ߡ������������(ͼ2)����������������(ͼ3)���Կ�����BY��������������������������������أ���������Խ�죬������������Խǿ����ȻBY����Ϊ����������������Fe2+Ϊ��Դ����, �����������������е��������ɻ���һ��[13]��

ͼ2 BY������������
Fig.2 Growth curve of BY strain

ͼ3 BY�������������������ͼ
Fig.3 Ferrous ion oxidation curves by BY strain
2.2.2 ����������
ͼ4��ʾΪBY������������˵������9K����������(5%����)21 d�ڣ�������pHֵ��ʱ��ı仯���ߡ�ͼ5��ʾΪ��Ӧ�����������У���9 d��15 d��21 dʱ����������ͼ���ɼ�����BY��������ֻ���������9K������������ǰ9 dʱ���pHֵ�����Ȳ��½����ɴ��Ʋ���Ѵ��ڶ����������У���18~21 d֮�䣬pHֵ�仯����ƽ������ʱ���Ѵ���ƽ�����У���21 d�Ժ�pHֵ����1.0���¡��Ӿ����������������ǰ9 d������������Ҫ���ڵ�9~15 d��������������9~15 d����������Ҫ���ڵ�15~21 d��������������������У���������������ʹ�������������ᣬ������������pHֵ�½����ɳ�ʼ��2.3����1.0��˵���þ����н�ǿ�����������ԡ�

ͼ4 BY�������������̵�pHֵ�ı仯
Fig.4 Variation of pH values in sulfur oxidation by BY strain

ͼ5 �������BY���ľ����仯
Fig.5 Population number of BY strain with sulfur substrate
2.3 ��������ø����
��������ø����Ϊ��A. f��������лϵͳ�Ĺؼ�ø[14-15]��Kusano��[16]��Ϊ�û�����1��237 bp�Ŀ����Ķ���(ORF)�����뺬90���������iro���ף����л�����37��������Ŀ�Ĥ�ź�����ͬʱ�������ȶ��ԣ���������������ø(���к�ΪCAF21316)��һ�����ʵ��ף�ͬʱ��ͨ�����бȽϺ��������֣�iro�������������λ����(HiPIPs)���кܸߵ�ͬԴ�ԡ����о�����Ϊ[14-16]����������ø�������Ǵ�Fe2+���ܵ��ӣ�Ϊ���Ӵ������еĵ�һ���������塣Ϊ�ˣ����о�����Kusano��[16]�����Ļ�������((�Ǽ����к�ΪE03451����СΪ500 bp)�������һ������ (YT1��YT2)������BY���Ļ�����DNAΪģ�������PCR��������¡������ⶨ�����г���Ϊ452 bp��Genbank�е����е�¼��ΪAY864809����ĿǰGenbank���������ݽ��жԱȣ������ʾ�������ĺ����������������� (�Ǽ����к�ΪDQ166839- DQ166841)����1�����������������������ȫһ�¡�
2.4 16S rRNA���з�����ϵͳ������
����PCR������������16S rRNA��Ȼ��¡�Ͳ��ⶨ�����г���Ϊ1 503 bp��Genbank�е����еǼǺ�ΪDQ062114����16S rRNA ����ͬԴ��Ϊ��������ϵͳ����������A.albertensis��Ϊ����Ⱥ����ͼ6��ʾ��BY������A.f��WJ13λ��ϵͳ������ͬһ����֧�У����ƶ�Ϊ99.66%����ĿǰGenbank�����м�¼������������ȣ��������ΪA.f������ATCC23270(�Ǽ����к�ΪAF465604)���ƶ�Ϊ99.93%���о�������A.f���ķֲ������Ϳ���������[17-18]���þ���ķ�����ΪA.f���Ķ����Ժ͵���ֲ��о��ṩ��һ���²��ϣ���Խ���ϸ�����Ŵ�����ѧ�о�����һ�������塣

ͼ6 BY�����16S rRNA����ϵͳ������
Fig.6 Phylogenetic tree derived from 16S rRNA sequence of BY strain
2.5 ����ʵ��
��ͭ���ϸ������ʵ������ݽ����ʾ���������4 d��Cu2+�Ľ����ٶȽϿ죬�����ǽ���Һ�����ԣ�������ѧ��Ӧ���¡��˺�ϸ����Cu2+�Ľ�������ռ������λ��ֱ��20 d��Cu2+�Ľ����ٶȱ����ȶ�����ʱ��仯��������20 d��Cu2+�Ľ������ֳ��ӿ�����ƣ���27 d��Cu2+Ũ�ȴﵽ60 mg/L(��ͼ7��ʾ)����ͭ��Ľ���ʵ����������BY�������һ���Ľ������������Ծ����һ��ѱ������Cu2+�Ľ������ʿ��ܽ�һ����ߡ�
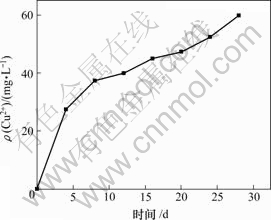
ͼ7 BY����Ի�ͭ���е�Cu2+�Ľ���
Fig.7 Copper extraction from chalcopyrite concentrate by BY strain
3 �� ��
a. �Ӹ�������������ˮ�з���õ���һ����������������˾�BY��Ϊ����������ϸ�����̸�״�������ֱ��������Ϊ(0.45��0.05) ��m��(1.5��0.4) ��m�����������¶�Ϊ25~30 �棬����pHֵΪ2.0��
b. �þ�����������������������������������������õ���������������������У������ˡ������Ƕ�BY������������������á��þ�����н�ǿ�������������ԡ���70 h�����������ʴﵽ99.5%���þ��껹���к�ǿ�����������ԣ���21 d������������pHֵ�ɳ�ʼ��2.3����1.0�������������Ժ����������ԵĻ������������������ء�BY������н�����ͭ�����������27 d��ϸ��������ͭ����Ũ�ȴﵽ60 mg/L��
c. 16S rRNAϵͳ�������Ĺ���������BY������A.f��WJ13λ��ϵͳ������ͬһ����֧�У����ƶ�Ϊ99.66%����ĿǰGenbank�����м�¼������������ȣ��������Ϊ����ATCC23270(�Ǽ����к�ΪAF465604)���ƶ�Ϊ99.93%����������ø����������ʾ�������ĺ��������뱨������(�Ǽ����к�ΪDQ166839- DQ166841)������1������IJ��죬��������������ȫһ�¡�
�ο����ף�
[1] Scott D J. The mineralogy of copper leaching: concentrates and heaps, copper��91[C]//Copper Hydrometallurgy Short Course. Santiago, 1995.
[2] Rawlings D E. Biomining: theory, microbes and industrial processes[M]. Berlin, New York: Springer�CVerlag, 1997: 229-245.
[3] YANG Song-rong, XIE Ji-yuan, QIU Guan-zhou, et al. Research and application of bioleaching and biooxidation technologies in China[J]. Minerals Engineering, 2002, 15: 361-363.
[4] Olson G J, Brierley J A, Brierley C L. Bioleaching review part B: Progress in bioleaching: applications of microbial processes by the minerals industries[J]. Applied Microbiology Biotechnology, 2003, 63: 249-257.
[5] Kelly D P, Wood A P. Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov. and Thermithiobacillus gen.nov. [J]. International Journal of Systematic and Evolutionary Microbiology, 2000, 50: 489-500.
[6] Rawlings D E. Heavy metal mining using microbes[J]. Annual Review Microbiology, 2002, 56: 65-91.
[7] SHI Shao-yuan, FANG Zhao-heng. Bioleaching of marmatite flotation concentrate by Acidithiobacillus ferrooxidans[J]. Hydrometallurgy, 2004, 75: 1-10.
[8] Brierley J A, Brierley C L. Present and future commercial applications of biohydrometallurgy[J]. Hydrometallurgy, 2001, 59: 233-239.
[9] Baker B J, Banfield J F. Microbial communities in acid mine drainage[J]. FEMS Microbiology Ecology, 2003, 44: 139-152.
[10] Silverman M P, Lundgren D C. Study on the chemoautotrophic iron bacterium Ferrobacillus ferrooxidans: I. An improved medium and harvesting procedure for securing high cell yield[J]. Journal of Bacteriology, 1959, 77: 642-647.
[11] ���ں�, �����, ������, ��. ����������˾��ľ�������о�[J]. ����ۺ�����, 2001(1): 19-23.
ZHANG Zai-hai, QIU Guan-zhou, HU Yue-hua, et al. The investigation of the colony isolation of Thiobacillus ferrooxidans[J]. Multipurpose Utilization of Mineral Resources, 2001(1): 19-23.
[12] Thompson J D, Gibson T J, Plewniak F, et al. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools[J]. Nucleic Acids Research, 1997, 24: 4876-4882.
[13] Ageeva S N, Kondrat��eva T F, Karavaiko G I. Phenotypic characteristics of Thiobacillus ferrooxidans strains[J]. Microbiology, 2001, 70(2): 186-194.
[14] Rawlings D E. The molecular genetics of Thiobacillus ferrooxidans and other mesophilic, acidophilic, chemolithotrophic, iron- or sulfur-oxidizing bacteria[J]. Hydrometallurgy, 2001, 59: 187-201.
[15] Yamanaka T, Fukumori Y. Molecular aspects of the electron transfer system which participates in the oxidation of ferrous ion by Thiobacillus ferrooxidans[J]. FEMS Microbiology Reviews, 1995, 17: 401-413.
[16] Kusano T, Takeshima T, Sugawara K, et al. Molecular cloning of the gene encoding Thiobacillus ferrooxidans Fe(I1) oxidase[J]. The Journal of Biological Chemistry, 1992, 267(16): 11242-11247.
[17] PENG Hong, YANG Yu, LI Xuan, et al. Sturcture analysis of 16S rRNA sequences of strains from Acidithiobacillus ferrooxidans[J]. Journal of Biochemistry and Molecular Biology, 2006, 39(2): 178-182.
[18] Karavaiko G I, Turova T P, Kondrateva T F, et al. Phylogenetic heterogeneity of the species Acidithiobacillus ferrooxidans[J]. Journal of Systematic and Evolutionary Microbiology, 2003, 53: 113-119.
�ո����ڣ�2006-11-18
������Ŀ�����Ҵ����о�Ⱥ���ѧ����������Ŀ(50621063)�����ҡ�973�������о���Ŀ(2004CB619201)
����飺�� ��(1972-)���У����ϳ�ɳ�ˣ����ڣ���������ѧ�о�
ͨѶ���ߣ����������У����ڣ��绰��0731-8836019��E-mail��HYH@mail.csu.edu.cn

